Le Centre National d’Agroécologie (CNA) est un projet porté par des acteurs de l’agroécologie qui y fédèrent leurs compétences et expertises : diffusion et échanges de savoirs, formation et accompagnement technique, outils d’évaluation et de mesure pour concevoir et piloter les systèmes agroécologiques. La raison d’être du CNA est de faire émerger des innovations basées sur les principes de l’agroécologie, au service de l’agriculture et des agriculteurs, pour aider à l’évolution et à la diffusion de leurs pratiques.
Produire durablement une nourriture de qualité
Sécuriser le revenu des agriculteurs
Couvrir et nourrir le sol pour protéger l’environnement
Atténuer le changement climatique
Stopper le gâchis d’énergie et ressources
Produire de la biodiversité
Limiter les pollutions systémiques
Créer du sens et de la solidarité
Copier la nature Toujours couvert/jamais travaillé
Recherche et formation …..sont des bases solides pour l’avenir
Produire et redonner du pouvoir aux agriculteurs ? C’est le pari de Lucien Séguy, « agronome », mais surtout « paysan ». Lucien Séguy dédiera sa vie à la fertilité et au « génie végétale ». Sans dogmatisme, il associera la plus grande diversité biologique possible et les techniques agro-industrielles de pointes, créant lui-même plusieurs variétés de semences de riz à haut-rendements.
Les militants de Solidarité & Progrès ont dénoncé une politique de la famine depuis la fin des accords de Bretton Woods (1971) et la publication du rapport « Halte à la croissance » (1972). Depuis lors, la dérégulation met tous les agriculteurs en compétition, accroissant les inégalités ; et l’idéologie décroissante des institutions financières s’oppose à l’équipement de l’agriculture, même là où elle est la seule ressource.
Est-il encore possible (écologiquement ? ) d’augmenter la productivité agricole ? Dans ce dossier, nous vous proposons de suivre le chercheur Lucien Séguy, à la découverte d’un monde longtemps passé inaperçu, celui des sols vivants. Nous verrons comment l’alliance des sols et des agriculteurs peut vaincre la pauvreté et la faim.
Biographie : Une vie au service de la science ; la science au service des paysans
Lucien Séguy en mission d’appui au Maroc. Source : Fert
Né en 1944 dans une famille paysanne de Dordogne, il est le seul de sa fratrie à poursuivre des études supérieures. Diplômé de l’Ecole Nationale Supérieur d’Agronomie de Toulouse, il se spécialise dans la science des sols (pédologie), puis part en service civique au Sénégal en 1967, développer la riziculture paysanne dans la région de la Casamance. Le Centre de coopération internationale en recherche en agronomique pour le développement (Cirad) lui confie sa première affectation professionnelle au Cameroun. Il y met en évidence l’importance de la gestion du sol pour lutter contre la pyriculariose, un champignon qui détruit les feuilles du riz.
1977 : repéré pour ses travaux sur les cultures de riz, il est appelé au Brésil. Débute sa véritable aventure scientifique. Il y travaillera jusqu’à sa retraite en 2009. En parallèle, il lance des missions d’appui aux paysans dans une dizaine de pays, dont Madagascar, la Côte d’Ivoire, le Gabon, le Cameroun, le Sénégal, la Tunisie, le Viêtnam, le Laos, le Cambodge, le Canada, la France métropolitaine et d’outre-mer, jusque dans les mois qui précèdent son décès en avril 2020.
Le texte qui suit est adapté librement du rapport de Lucien Séguy, son ami Serge Bouzinac, et leurs collègues de la recherche brésilienne : « La symphonie inachevée du Semis Direct dans le Brésil central », rédigé en 2008 pour le Cirad et l’Embrapa (la Société Brésilienne de Recherche Agricole).
Les effets spectaculaires de l’érosion en Europe. Source : agriculture-de-conservation.com / C. Henricot
Lorsque Lucien Séguy arrive au Brésil, il rencontre des paysans ruinés par la dégradation du sol. En cause, le labour.
Le labour est pourtant une méthode qui a fait ses preuves en agriculture. Depuis sa découverte dans le croissant fertile, il y a 10 000 ans, plusieurs civilisations agraires y ont eu recours, de la Mésopotamie à l’Europe et l’Asie. Le labour a permis à des centaines de générations de se nourrir, jusqu’à nous.
Il empêche l’installation des mauvaises herbes et des maladies du sol. Mais exige d’énormes quantités de labeur des paysans. Avec un risque de désertification, particulièrement rapide sous les tropiques.
En climat tropical, les sols ont un aspect minéral, comme la terre de remblai peu fertile de nos terrains vagues. Seuls les cinq premiers centimètres de surface sont noirs et fertiles, comme le terreau ou le compost.
Cette mince couche ne peut s’épaissir, car l’humidité et la température tropicale accélèrent tous les processus du vivant : la croissance des plantes cultivées, comme la décomposition de la matière morte au sol. La « matière organique », noire et fertile, est donc fragile sous ces latitudes.
Dans ce contexte, la technique du labour, qui apporte beaucoup d’oxygène au sol, accélère la décomposition de la matière organique, par les microorganismes du sol. C’est « l’érosion invisible ». De plus, mis à nu et cassé en mottes, le sol est emporté dans les fleuves par les puissantes pluies tropicales. C’est « l’érosion visible ».
En forêt, rien ne se perd !
Tout comme Louis Pasteur, Lucien Séguy a passé une partie de sa jeunesse à peindre, exerçant et développant ses talents d’observateur. En grand scientifique, il interroge sans cesse la nature : « Mon maître, mon juge, mon centre d’inspiration principal, c’est la nature, sous sa complexité la plus grande. Vous avez une question à poser ? Posez-la à la nature. Et elle vous répondra ! Il faut simplement faire quelques manip’ pour qu’elle vous réponde. Et le faire de manière scientifique pour comprendre pourquoi cette réponse ; ce qu’elle veut dire exactement. »
Le but de Séguy est d’inventer une agriculture inspirée de la fertilité naturelle des forêts. Il fonde les principes de cette nouvelle agriculture sur quelques observations.
Lucien Séguy note le rôle essentiel de la « matière morte » dans la fertilité des forêts tropicales. Les « éléments fertilisants » qui s’enfonceraient dans le « sol minéral » pouvant malgré tout être recyclés par les racines profondes.
– Une première observation : Le sol forestier n’a pas besoin d’être labouré pour être meuble. Les organismes du sol forestier et les racines des plantes brassent la matière et produisent la porosité nécessaire à la présence raisonnable d’oxygène et d’eau dans le sol forestier.
1er principe : Lucien Séguy propose donc de semer directement les graines dans le sol, sans labour. L’idée n’est pas nouvelle. Ce semis dit « direct » avait permis aux civilisations précolombiennes de prospérer. La méthode en fut oublié, avec leur effondrement. Cette technique est redécouverte aux Etats-Unis dans les années 60, pour parer aux tempêtes de sable ; suite au « Dust Bowl » des années 30, lors de la grande dépression, qui a profondément meurtri le pays, comme l’illustre le film Les raisins de la colère.
C’est le glyphosate, bêtement diabolisé par des écologistes radicaux, qui a permis aux Etats-Unis d’abandonner le labour, donc d’éviter le désert… Le glyphosate agit en bloquant la photosynthèse des mauvaises herbes, rendant caduc l’archaïque labour. À ce titre, Lucien Séguy propose avec humour que soit décerné « le prix Nobel de la paix à Monsanto, pour sa contribution à la préservation des sols ».
Mais – attention – Lucien Séguy est lucide quant au risque de destruction de l’agriculture par les biotechnologies, quand elles sont développées dans un but mercantile. Au sujet de l’herbicide de Monsanto et des OGM, il nous dit : « la productivité n’a pas augmenté depuis leur adoption massive [alors] que les doses d’herbicide sont plutôt en croissance pour contrôler les [plantes] devenues résistantes au glyphosate. » Avoir recours à de plus grands volumes, inutilement… le contraire de l’écologie !
En Semis Direct, le sol n’est pas retourné : à l’avant du tracteur, un « rolofaca » ou « rouleau hacheur » couche le couvert végétal au sol ; à l’arrière, le semoir sème directement les graines sous cette litière. Source : paysan-breton.fr / Toma Swan
– Lucien Séguy fait une deuxième observation : Les sols forestiers sont couverts en permanence d’une litière végétale. Sous cette litière, les premiers centimètres du sol sont le siège de la fertilité de la forêt tropicale. En profondeur, les éléments nutritifs se font beaucoup plus rares !
2nd principe : Pour avoir un sol aéré, riche en nutriments et protégé contre les pluies diluviennes, Lucien Séguy comprend qu’il doit restituer une partie de la culture au sol. L’enjeu est notamment de nourrir les vers de terre et leur cortège d’êtres vivants. Il choisit de recourir à de « puissantes biomasses » pour couvrir le sol et le régénérer en matière organique, qui confère sa fertilité au sol.
Semis direct dans un couvert de sorgho en Europe. La densité et la vigueur du sorgho en font une excellente « plante géante » de couverture, pour contenir les autres plantes et nourrir le sol. Source : agriculture-de-conservation / Cécile Waligora.
– Une ultime observation permet à cet « ingénieur du végétal » d’identifier la complémentarité des plantes dans les systèmes forestiers : toutes ne prospectent pas l’eau aux mêmes profondeurs ; et certaines plantes peuvent arracher des éléments minéraux au sol, pour les mettre à disposition des plantes de la culture suivante.
3ème principe : Lucien Séguy commence donc un travail qui est au cœur de ce qu’il nomme sa « symphonie inachevée » : Il recherche les meilleures successions et associations végétales, permettant de réduire les dépendances aux fertilisants et aux pesticides, et de maximiser les revenus des agriculteurs.
En rupture avec la monoculture, il propose : « L’incorporation de biomasses de couverture encore plus puissantes et plus diversifiées pour que les couverts végétaux des SCV gagnent en multifonctionnalité gratuite. »
Entre deux cultures commerciales, des couverts végétaux sont semés pour réorienter la biologie, la chimie et la structure du sol. Ici, un mélange de tournesol, phacélie et pois. Source : Ver de terre production
Lucien Séguy appelle sa méthode : « Semis sous Couvert Végétal » (SCV).
Avec son binôme Serge Bouzinac et ses partenaires de la recherche et développement brésiliens, Séguy met en place une méthodologie de « recherche-action » sur « système de cultures pérennes » : le chercheur fait ses essais avec, pour et chez les agriculteurs, sur plusieurs années, pour adapter ses travaux aux contraintes de la production.
Pour ces travaux, Lucien Séguy est nommé docteur « Honoris causa » de l’université de Ponta Grossa, une des meilleures universités agricoles du monde, et “Grand Citoyen” par l’Assemblée Législative de l’Etat du Mato Grosso. Mais le paysan-chercheur tient à rendre hommage aux efforts des agriculteurs brésiliens et des acteurs institutionnels :
« Le Brésil a montré au monde sa capacité à développer en moins de 30 ans une agriculture de conservation que le monde entier admire et lui envie, même si elle est largement encore perfectible comme l’ont démontré nos travaux sur les innovations SCV inspirées du fonctionnement de l’écosystème forestier. »
Ajoutons que, bien que fortement mécanisés, les agricultures du Brésil, de l’Argentine et des Etats-Unis restent peu productives, car peu irriguées : dans la région du Mato Grosso entre 2001 et 2007, les rendements par hectare sont en moyenne : de 3,5 tonnes pour le coton, 2,9 tonnes pour le soja et 3,4 tonnes pour le maïs. En France, le blé produit 7 tonnes par hectare et le maïs 9 tonnes. Un ami et disciple de Lucien Séguy, Christian Abadie, produit même 14 tonnes de maïs dans le sud-ouest de la France !
Pourquoi les agriculteurs des Etats-Unis, de l’Argentine et du Brésil acceptent-ils de produire si peu par hectare ? La réponse est en partie économique.
Regardons plutôt, pour le moment, les résultats de la méthode du « Semis sous Couvert Végétal » de Lucien Séguy.
Bilan des « Semis sous Couvert Végétal »
Amélioration du statut social des paysans, accomplissements économiques et environnementaux divers ; les résultats présentés ci-dessous sont non exhaustifs, mais sont l’occasion d’illustrer les problématiques de l’agriculture.
Résultats du groupe agro-industrielMAEDA : Augmentation de la productivité du soja de 25%, et du coton de 45%. Soit une productivité respectable de près de 4 tonnes de soja par hectare et 5 tonnes de coton. Passage à 3 cultures sur 2 ans au lieu de 2. Les marges ont été multipliées par 3. Le nombre de machines agricoles a été réduit de moitié. Le nombre de prestataires de services a chuté de 71%. Et la consommation de carburant a diminué de 70%. Produire plus avec moins !
Des engrais renouvelables : Le manque d’azote et de phosphore dans le sol est ce qui limite le plus la productivité des champs. L’azote constitue 78% de l’air que nous respirons. Il est obtenu industriellement par la réaction du gaz naturel avec l’air. Quant au phosphore, il est aujourd’hui extrait des carrières de « guano », sites de déjections des oiseaux marins, ressource peu renouvelable.
A titre d’exemple, Lucien Séguy introduit la plante sauvage Stylosanthes, pour sa capacité à fixer l’azote atmosphérique et à mobiliser le phosphore du sol. Cet exploit naturel est dû à des symbioses racinaires, avec des bactéries pour l’azote, et avec des champignons pour le phosphore.
En plus de la Stylosanthes, il choisit la Brachiaria, pour ses racines profondes qui récupèrent les nutriments perdus, de même que pour la densité de son couvert au sol et pour sa forte productivité. Lucien Séguy sème ces deux plantes entre les pieds du maïs qui précède la culture de soja. Il en tire une hausse de productivité pour le maïs et le soja, et s’en sert comme fourrage pour l’élevage.
Une réduction des dégâts : Sur soja, cette fertilité organique permet de réduire le nombre de traitements fongicides des 2/3, voire totalement, et d’obtenir des grains de 1 à 3% plus riches en protéines.
Sur coton, en réduisant de 30 à 50% les apports chimiques d’azote et de potassium, en simple complément de la fertilité naturelle du sol, les dégâts liés au ravageur sont nettement plus faibles. Les passages de pesticides peuvent alors être réduits d’1/3.
Une auto-épuration naturelle : Le sol vivant est un « biodigesteur » des résidus végétaux, mais aussi des pesticides, qu’il réduit en molécules de plus en plus simples. Plus il y a de matière végétale au sol, plus les pesticides restent piégés longtemps dans ce biodigesteur, et donc meilleure est la destruction de ces polluants.
En culture de coton, qui exige souvent de nombreux traitements chimiques, les Semis sous Couvert Végétal (SCV) permettent de réduire le volume d’insecticides de près d’un tiers et le nombre de molécules différentes de 18 à 5. Le volume de fongicides (contre les champignons) est réduit de moitié. La fonction auto-épuratrice des sols vivants permet de réduire sous le seuil de détection l’ensemble des 150 molécules et sous-produits polluants recherchés.
Une valorisation du territoire : Vous voulez sauver la forêt amazonienne ? 1/5ème des surfaces déboisées de l’Amazonie sont à l’abandon, car dégradés par le travail du sol. Les SCV peuvent régénérer ces 16,5 millions d’hectares :
« Les SCV les plus puissants permettent, en 3 à 5 ans, de retrouver les teneurs en matière organique et les caractéristiques de distribution des tailles d’agrégats des sols originels sous forêt. »
Autres sols valorisables : les savanes arbustives du Brésil, appelées Cerrados, réputés incultes. Ces terres, qui rappellent nos garrigues méditerranéennes, offrent plus de 50 millions d’hectares faciles à cultiver en SCV.
Avec la régénération des sols forestiers détruits et la valorisation des sols de savane incultes, Lucien Séguy se réjouit que le Brésil puisse doubler ses surfaces de production sans détruire la forêt amazonienne !
Le bocage Normand. Les haies et les arbres créent un microclimat à l’échelle des parcelles. Auteur : Bournagain.
Enfin, l’eau étant un élément essentiel à la vie, Lucien Séguy recommande de maintenir un maillage continu de végétation native de 20 à 50 mètres autour des parcelles, plutôt que des îlots forestiers. Cette « forme géométrique du défrichement » permet d’éviter l’ « effet de poêle surchauffée qui engendre des courants ascendants d’air chaud qui rejettent les pluies vers les forêts galeries à la périphérie ».
Une symphonie “inachevée”
Comme expliqué précédemment, les agricultures de hautes technologies du Brésil, de l’Argentine et des Etats-Unis pourraient assez facilement produire 1 tonne de plus par hectare. Lucien Séguy déplore le manque de formation technique et pointe aussi des raisons économiques.
À l’image des agriculteurs des Etats-Unis, les agriculteurs du Brésil ont adopté le non-labour, poussés par l’érosion des sols et par les prix très bas payés aux productions agricoles. Le non-labour leur permet de réduire leurs dépenses en matériel et essence, et apporte de la durabilité à leurs exploitations.
Au Brésil Central, où 95% des agriculteurs pratiquent le Semis Direct (1er principe : non-labour), le Semis sous Couvert Végétal (2ème & 3ème principes : une végétation puissante et diversifiée) n’est en réalité pratiqué que sur 30% des champs de soja et 15% des champs de maïs.
Ainsi, depuis les années 2000, étant insuffisamment régénérés, les sols du Brésil Central se compactent et un travail superficiel du sol redevient nécessaire. La méthode alors utilisée est le « discage » des 5 premiers centimètres, ce qui constitue une régression vers le labour et la chimie, des surcoûts et une perte de durabilité.
C’est que produire les couverts à aussi un coût ! Les systèmes de SCV nécessitent des investissements de 11% à 29% supérieurs aux simples systèmes sans labour. Hélas, en plus des aléas naturels, le libéralisme rend les prix payés aux producteurs extrêmement fluctuants et généralement plutôt bas. Ainsi, il est probable que les lendemains incertains découragent l’épargne dans le sol.
La formation professionnelle et des marchés organisés apparaissent comme les deux leviers pour qui veut bâtir une politique du sol vivant.
A la frontière des travaux de Séguy, plusieurs disciplines peuvent être convoquées :
Une myriade de naturalistes (dont les spécialisations sont en proportion de la créativité de la Nature) et d’écologues (spécialisés, eux, dans l’étude des interactions naturels) : mobilisés auprès des agronomes (à recruter également !) pour rechercher de nouvelles plantes de couvertures et de nouvelles stratégies d’assemblage du vivant.
Encore une question de moyen ! L’étude des systèmes naturels nécessite des moyens logistiques que l’approche statistique et la modélisation informatique, généralement privilégiées aujourd’hui en écologie scientifique car moins onéreuses, ne devrait qu’appuyer !
Comme pour les plantes que nous mangeons et qui ont été sélectionnées pour être productives, les sélectionneurs peuvent améliorer les caractéristiques des plantes de couverture : Le sorgho blanc sans tanins à haute teneur en protéines, pour farines, bière, papier et amylose ; le sésame qui contient de 50 à 55% d’huile de qualité supérieure, pour les cosmétiques et l’aviation ; le sarrasin, pour les farines sans gluten, …
L’acclimatation de nouvelles plantes de cultures est aussi un moyen d’améliorer la santé du sol. Par exemple, l’intégration du riz pluvial dans les rotations avec les cultures de soja et coton permettrait de lutter contre les maladies à champignon. Pour rendre cette culture de riz pluvial attractive, le défi est actuellement de mettre au point des variétés à hauts rendements.
Quant aux biologistes et aux généticiens, leurs connaissances à l’échelle cellulaire et moléculaire est une aubaine pour l’étude du sol vivant :
Le séquençage génomique, d’abord, pour l’étude de la diversité microbienne. Champignons et bactéries cachent d’innombrables possibilités de symbioses et d’effets antibiotiques naturels. Il s’agit de tirer parti du haut potentiel des sols vivants !
La recherche dans les OGM aussi, à condition d’être au service de l’agronomie, seule discipline globale des champs : « Les SCV offrent une biodiversité fonctionnelle très efficace […] dans un tel contexte, les OGM pourraient être des auxiliaires précieux, légitimes et incontestablement valorisés pour seulement compléter les services écosystémiques majeurs des SCV. »
Nourrir les BRICS
Lucien Séguy, qui dénonce volontiers le conservatisme français, ne s’étend pas sur la politique du Brésil. Tentons, par nous-même, de comprendre cette très jeune puissance agricole :
L’Embrapa, entreprise d’Etat, a acclimaté le blé, le soja, le maïs et le coton, aux sols et aux climats brésiliens. Son existence depuis 1972 inscrit le Brésil dans une démarche scientifique à long terme et ouvre la possibilité de transferts de compétences avec l’Afrique.
Tracés envisagés du chemin de fer “bi-océanique”.
Bien que le Brésil ait su défendre ses intérêts agricoles devant l’Organisation Mondiale du Commerce, le pays tente dès 2003 de remettre en cause les règles arbitraires imposées par les institutions dévoyées de Bretton Woods. En 2014, au sommet de Fortaleza, est lancée la Nouvelle Banque de Développement des BRICS (Brésil, Russie, Inde, Chine et Afrique du Sud).
Il s’agit pour les pays émergents de réduire leurs coûts de production, non par le dumping, mais par l’apport technologique et l’aménagement du territoire. Cette politique est aussi connue sous le nom de « Nouvelles Routes de la Soie ». Un exemple de coopération agricole entre la Chine et le Brésil : la construction de la voie ferrée bi-océanique, traversant l’Amérique latine d’Est en Ouest. Projet gagnant/gagnant, puisqu’il désenclave l’Amazonie et évite le passage par le canal de Panama pour fournir la Chine en denrées agricoles.
En France, les réseaux de Lucien Séguy ont débuté 30 ans après le bio, mais 15% des surfaces agricoles françaises sont déjà cultivées en Semis-direct, soit le double de l’agriculture biologique. En dehors des radars de la plupart des médias.
Bien sûr, le bio nourrit aussi les gens et doit être soutenu à ce titre. Notons seulement ses limites pour mieux nous détacher de l’agribashing : Une productivité moindre et un sol moins riche en vie.
Apprécions aussi les vrais mérites du bio : une production fraîche, car locale, et des circuits-courts, pour que la valeur aille aux producteurs.
Lucien Séguy entouré de Français passionnés. Source : festival Paysage in Marciac, 2018.
Les réseaux français de Lucien Séguy tentent toutes les dénominations pour se faire connaître du public : agriculture de conservation, agriculture de régénération, agriculture sur sol vivant. Et le ver de terre comme symbole de la méthode.
Ces agriculteurs sont présents dans tous les syndicats. Et la Coordination Rurale, second syndicat agricole de France, entre la FNSEA et la Confédération Paysanne, organise depuis 25 ans des festivals de non-labour.
Un ministre, le socialiste Stéphane Le Foll, a rencontré les leaders de ces mouvements en France, mais son cabinet n’a rien trouvé de mieux à leur proposer qu’une tribune à la COP21. Car la priorité du ministère est d’atteindre la neutralité carbone de notre agriculture, d’ici à 2050. Notre article « Ces milliardaires « écolos » qui veulent nous affamer » dénonce cette politique agricole malthusienne.
Une rencontre fortuite nous avait d’ailleurs permis d’interroger le ministre sur la détresse des agriculteurs et les prix trop bas. Nous lui demandions de mettre en place une limitation des importations et un stockage des surplus pour hausser les prix. Réponse du ministre : « Nous ne sommes pas communistes ! » Ce fut l’occasion pour nous d’évoquer la méthode des trente glorieuses : « Monsieur le ministre, De Gaulle non plus ; pourtant, c’était sa politique ».
Stéphane Le Foll a tout de même soutenu la création de « ver de terre production ». Cette chaîne de diffusion de contenus sur Youtube est animée par des agriculteurs. Mais les médias traditionnels restent en roue libre… et le public avec eux. Le pays a-t-il besoin d’une chaîne de télévision publique, qui soit dirigée par les instituts scientifiques ?
Parmi les mesures pour encourager l’agriculture sur sol vivant, rappelons l’importance d’autoriser le glyphosate pour maîtriser les couverts végétaux.
Enfin, l’hexagone gagnerait beaucoup à mettre en place une taxe sur les importations de soja. Elle inciterait le retour de la production de légumineuses en France, comme durant l’année 1973 (lorsque les Américains ont organisé un embargo du soja contre l’Europe).
Qu’est-ce qu’une légumineuse ? Une légumineuse est une plante qui, comme le pois, la lentille ou le soja, est capable de fixer l’azote présent dans l’air. Elles permettent de fertiliser un champ pour la culture suivante. Les légumineuses servent à diversifier les cultures pour limiter l’installation de maladies du sol. Et leurs graines, riches en protéines, augmentent l’autonomie de l’élevage. Cette taxe est donc une mesure de « protectionnisme éducateur », dans la tradition de l’économiste Friedrich List.
Le Brésil a prouvé qu’il est capable d’entendre cet argument, à condition que son droit au développement soit pris en compte. Retrouvons l’état d’esprit des coopérants, comme Lucien Séguy.
Antoine Beils, mai 2021
Annexe I : De l’importance de la productivité « physique » pour nourrir le monde
L’économiste et homme politique américain Lyndon Larouche a montré la relation entre d’une part, la productivité par travailleur, unité de surface et volume de matières utilisées (les 3 paramètres de la productivité physique) contraints par les handicaps naturels d’un territoire ; et d’autre part, le potentiel démographique de ce même territoire. Dit autrement : Le nombre d’humains que la Terre peut porter n’est pas déterminé par les ressources naturelles, mais par l’efficacité de leur utilisation.
Examinons la productivité physique de l’agriculture.
L’agriculture raisonnée, l’agriculture de précision et l’agroécologie permettent toutes trois de produire autant, avec de moindres volumes de produits chimiques. Ce qui n’est pas forcément le cas du bio, qui a globalement recours à de grands volumes de pesticides « naturels ».
Même constat, avec la faible production du bio par hectare et par travailleur ; bien que l’émergence de pratiques agroécologiques (lorsqu’elles sont compatibles avec le cahier des charges du bio) permettent quelques améliorations.
L’Organisation des Nations Unies (ONU) et sa branche pour l’Alimentation et l’Agriculture (FAO) soutiennent cette productivité physique. Elles parlent d’ailleurs d‘ « agriculture écologiquement intensive » ; mais sans moyens financiers à long terme pour promouvoir les semences à hauts rendements, la mécanisation et les autres moyens logistiques du stockage, du transport et de la chaîne du froid. D’ailleurs, le Programme Alimentaire Mondial des Nations Unies cherche encore 5,5 milliards de dollars, pour l’aide alimentaire d’urgence en 2021.
L’écologie, c’est quand les gros mangent les petits ?
Mais si les prix tombent trop, les agriculteurs renoncent à lutter contre les maladies. Au risque de perdre la production…
C’est aussi le risque pris par le bio, qui, en renonçant à la chimie de synthèse, produit globalement un tiers moins de nourriture que l’agriculture conventionnelle à l’hectare, mais est mieux valorisé commercialement.
La page du productivisme, tant décrié encore, a donc été tournée de force ; bien que les extrémistes verts ne s’en satisfassent pas et exigent l’abandon total des équipements motorisés et de la chimie.
Et le pouvoir de nourrir change de main :
1/ Dépossession des agriculteurs. Puisqu’il faut acquérir du foncier, il faut s’endetter au-delà de ce qui est remboursable. Un problème pour la transmission aux jeunes, mais pas un problème pour le rachat par des fonds financiers.
2/ Concentration de l’agro-industrie. À l’image des abattoirs géants, qui imposent leurs conditions commerciales aux éleveurs.
Annexe II : Puit de carbone & pompe à fric
En climat tropical, le Semis Direct sous puissante biomasse permet de produire de 23 à 32 tonnes de matière sèche, aérienne et racinaire, par hectare. Or, cette matière sèche est faite pour moitié de carbone. Si l’intégralité de cette biomasse est restituée au sol, alors 900 kilos à 2 tonnes de carbone sont séquestrés dans ce sol chaque année, et jusqu’à une dizaine d’années de suite.
Comme développé dans le dossier « Le New Deal vert : sortir du piège de la finance verte », la finance verte tente de mettre les États sous tutelle au nom du climat. Pour l’agriculture, aussi, accepter l’agenda “zéro carbone” est dangereux, car cet agenda mène à l’abandon des énergies fossiles, de la fertilisation azotée et de l’élevage, pourtant nécessaires pour nourrir correctement 9 milliards d’êtres humains en 2050.
Annexe III : Une plongée dans le sol vivant
Voir la vidéo : « Lucien Séguy, semis direct sur 20 millions d’hectares ». École Nationale Supérieure Agronomique de Toulouse (ENSAT).
Arnaud Beils says:Merci pour cet excellent article où j’ai appris beaucoup de choses sur l’intérêt du Semis direct sur Couvert Végétal. Je note notamment la capacité de certaines plantes à aller chercher les minéraux en profondeur. A noter également l’importance méconnus de cette pratique en France (15% de notre production !) Peut-être le semis sous couvert végétal permet-il aussi de réaliser des économies d’eau grâce au fait que le couvert végétal conserve l’humidité contrairement à de la terre nue ?Je souhaiterais te poser quelques questions pour comprendre ce que tu as écrit à fond. 1/ Pourquoi le Brésil n’arrive t’il pas au même rendement que la France ? Tu précises qu’ils sont fortement mécanisés et qu’en 2007, nous (la France) arrivons au double du rendement par hectare. Hypothèse, le sol n’est pas encore assez généré ?2/ L’équivalent du semis direct sous couvert végétal appliqué à de faibles surfaces (- de 1 hectare) est il le maraichage sur sol vivant ? Autrement, quels outils utiliser pour produire en SCV sur une faible surface ?3/ peut-on dire que l’augmentation du CO2 dans l’atmosphère (infime précisons le tout de même) est en parti dût au fait que l’on ne restitue pas la matière organique à la terre ? (d’où un argument supplémentaire en faveur du SCV pour capter le CO2)Encore merci pour cet apport intellectuel. Militons pour que la recherche agronomique ait les moyens d’augmenter notre potentiel démographique. Vive l’Humain explorateur du vivant !Reply
antoinebeils says:Merci à toi l’ami,Pour répondre rapidement : 1/ Comment doubler ou tripler la production : – Le Brésil est un pays aux climats contrastés. Le Mato Grosso par exemple, où a travaillé Séguy, est sec une partie de l’année. Il faut donc investir dans l’irrigation. – Une vraie régénération des sols permet les symbioses racinaires avec la vie du sol, pour valoriser la fertilité, donc une meilleur expression des potentialités de la plante. – Précisions : le soja étant 2 à 3 fois plus riche en protéines que le maïs, les faibles rendements sont compensés.2/ Comment appliquer l’ “agriculture sur sol vivant” sur une surface d’1 hectare : – La chaîne youtube “Ver de Terre Production”, animée par des agriculteurs et des chercheurs, permet de se mettre à jour de “l’état de l’art”, du jardin nourricier aux grandes cultures, en passant par l’élevage, l’arboriculture et le maraichage. 😉3/ La fixation du carbone, un argument pour les sols vivants ? Oui, mais… Le paradigme politique “neutralité carbone en 2050” est dangereux, car il implique la suppression de la fertilisation azoté et le démantèlement de l’élevage, comme l’exigent certains financiers fanatiques. Parlons plutôt de “l’agriculture sur sol vivant” comme d’une méthode pour nourrir le monde, bien nourrir, tout le monde !
Il s’agit d’une glycoprotéine hydrophobe tolérant la chaleur que l’on retrouve à la surface des spores et des mycéliums mycorhiziens. La production de cette substance atteint son maximum chez les mycéliums sénescents. Cette substance à décomposition lente comporterait le tiers du carbone séquestré dans les sols de planète. Sa principale fonction porte sur la stabilisation des agrégats, à la façon d’une colle regroupant l’argile, le limon et le sable fin, avec des effets majeurs sur les propriétés physiques des sols. Ce grumelage assure une meilleure pénétration de l’eau, réduisant les écoulements de surface, de meilleurs échanges gazeux et une rétention de l’eau et des minéraux, notamment le potassium. On peut dire que la glomaline est très étroitement liée à la fertilité des sols. On peut observer la glomaline par immunofluorescence sur les mycéliums, les spores et les agrégats de sol.
Malgré l’utilisation généralisée du glyphosate pour lutter contre les mauvaises herbes en agriculture, des questions demeurent quant à l’effet de l’herbicide sur les communautés microbiennes du sol. La littérature scientifique existante contient des résultats contradictoires, allant de l’absence d’effet observable du glyphosate à l’enrichissement en agents pathogènes agricoles tels que
Fusarium spp. Nous avons mené une étude approfondie sur le terrain pour comparer les communautés microbiennes présentes sur les racines des plantes ayant reçu une application foliaire de glyphosate avec celles des plantes adjacentes qui n’en ont pas reçu. L’étude de deux ans a été menée à Beltsville, MD, et Stoneville, MS, avec des cultures de maïs et de soja cultivées dans divers systèmes agricoles biologiques et conventionnels. En séquençant des amplicons de métabarcodes environnementaux, les communautés procaryotes et fongiques ont été décrites, ainsi que les propriétés chimiques et physiques du sol. Des sections de racines de maïs et de soja ont été étalées pour détecter la présence d’agents pathogènes végétaux. La géographie, le système agricole et la saison étaient des facteurs importants déterminant la composition des communautés fongiques et procaryotes. Les parcelles traitées au glyphosate ne différaient pas des parcelles non traitées en termes de composition globale de la communauté microbienne après contrôle des autres facteurs. Nous n’avons pas détecté d’effet du traitement au glyphosate sur l’abondance relative d’organismes tels que
Fusarium spp.
IMPORTANCE Accroître l’efficacité des systèmes de production alimentaire tout en réduisant les effets négatifs sur l’environnement reste un défi sociétal clé pour répondre avec succès aux besoins d’une population mondiale croissante. L’herbicide glyphosate est devenu un composant presque omniprésent de la production agricole à travers le monde, permettant une adoption croissante de l’agriculture sans labour. Malgré cette utilisation généralisée, de nombreux débats subsistent sur les conséquences de l’exposition au glyphosate. Dans cet article, nous examinons l’effet du glyphosate sur les communautés microbiennes du sol associées aux racines des cultures résistantes au glyphosate. À l’aide de techniques de métabarcoding, nous avons évalué les communautés procaryotes et fongiques à partir d’échantillons de sols agricoles (
n = 768). Aucun effet du glyphosate n’a été constaté sur les communautés microbiennes du sol associées aux variétés de maïs et de soja résistantes au glyphosate dans divers systèmes agricoles.
INTRODUCTION
Les microbes associés aux cultures agricoles affectent de multiples dimensions de la santé des plantes. Ils peuvent jouer des rôles importants liés à la physiologie des plantes, tels que l’acquisition de nutriments (
3 ), en plus d’aider à la défense contre les facteurs de stress biotiques (
4 ) ou d’agir en tant qu’agents pathogènes importants. Malgré cette importance pour la santé des plantes, ce n’est que récemment que la gestion de la diversité microbienne est considérée comme une possibilité réaliste d’augmentation durable de la productivité des cultures nécessaire pour répondre à la demande alimentaire face à la croissance de la population humaine et au changement climatique (
5 – 7 ). L’intensification de l’agriculture moderne a été motivée par l’utilisation de pesticides, d’engrais et d’autres amendements connus pour affecter les communautés microbiennes du sol (
8 ). Cependant, de nombreuses études ne disposent pas de la réplication spatiale et temporelle nécessaire à la rigueur statistique (
9 ). Une meilleure compréhension de la façon dont les systèmes agricoles (y compris les cultures) et la géographie interagissent pour façonner les communautés microbiennes est nécessaire afin de tirer parti des microbiomes agricoles pour la sécurité alimentaire (
10 ).L’introduction de cultures génétiquement modifiées résistantes au glyphosate (GR) a transformé les agroécosystèmes dans une grande partie du monde en augmentant l’adoption d’une agriculture sans labour et avec un labour réduit où les mauvaises herbes sont contrôlées chimiquement (
12 ). Les systèmes agricoles sans labour améliorent la structure du sol et la rétention des éléments nutritifs en réduisant l’érosion tout en réduisant les dépenses et la consommation de combustibles fossiles associés au fonctionnement des machines. Les communautés microbiennes dans les sols sans labour sont généralement plus diversifiées que celles dans les systèmes labourés en raison de l’augmentation de l’hétérogénéité des niches (
14 ).Le glyphosate interrompt la voie de biosynthèse du shikimate (
15 ), responsable de la production d’acides aminés aromatiques et d’autres composants clés du métabolisme cellulaire. La voie shikimate se retrouve chez les bactéries, les champignons, les algues, les plantes et certains protozoaires, mais pas chez les animaux. Le glyphosate se lie de manière compétitive à l’enzyme 5-énolpyruvylshikimate 3-phosphate synthase (EPSPS) par rapport au phosphoénolpyruvate, et il est mortel pour la plupart des espèces de plantes et une grande proportion de champignons (
16 ). Cependant, certains microbes sont résistants au glyphosate en raison du métabolisme rapide du glyphosate ou d’une forme GR du gène codant pour l’EPSPS (
17 ). Une fois cette voie de biosynthèse bloquée, les plantes meurent à cause d’une perturbation métabolique. Même à des taux d’application sublétaux, le glyphosate peut affaiblir suffisamment les défenses pathogènes d’une plante pour que les agents pathogènes puissent infecter et tuer la plante (
19 ). En l’absence d’agent pathogène, la plante peut avoir un aspect rabougri pendant quelques semaines, puis se rétablir.Le glyphosate est un herbicide appliqué foliaire qui se déplace rapidement du feuillage vers le reste de la plante, y compris les racines (
20 ). Les plantes peuvent exsuder du glyphosate de leurs racines dans les 24 heures suivant l’application foliaire (
22 ). Le glyphosate se lie fortement à certains composants du sol, étant presque immobilisé dans la plupart des types de sol (
23 ). Sa liaison étroite avec le sol contribue à sa faible phytotoxicité pour les plantes en tant qu’herbicide appliqué au sol. L’exsudation épisodique de glyphosate par les racines peut avoir des effets indirects sur la communauté microbienne du sol, et ces changements peuvent être importants pour la durabilité à long terme des agroécosystèmes. Cependant, les changements dans la communauté microbienne sont difficiles à détecter étant donné les effets simultanés de la saisonnalité, de l’évolution des espèces cultivées et du type de sol.Bien que l’agriculture sans labour présente des avantages évidents, les rapports diffèrent quant à l’effet du glyphosate sur la diversité microbienne. Des inquiétudes ont été soulevées concernant l’augmentation des charges pathogènes et la suppression des organismes bénéfiques associées à l’utilisation du glyphosate (
25 ). Il existe plusieurs mécanismes par lesquels le glyphosate pourrait enrichir le sol en agents pathogènes des plantes, comme suit : (i) les agents pathogènes pourraient attaquer les mauvaises herbes sensibles au glyphosate qui succombent à l’herbicide, dont la biomasse mourante agit alors comme refuge pour une infestation ultérieure des cultures (pont vert ); (ii) les agents pathogènes pourraient « prendre pied » dans une plante résistante au glyphosate en raison d’une réponse immunitaire réduite due à des altérations de la voie du shikimate, entraînant une infection non mortelle tout en permettant à l’agent pathogène de se propager ; et (iii) l’élimination des taxons microbiens sensibles pourrait également entraîner une réduction de la concurrence pour l’espace des niches racinaires, permettant ainsi aux agents pathogènes d’accéder aux tissus végétaux. Un examen de toutes les cultures GR par Hammerschmidt (
19 ) a déterminé qu’il n’existe aucune preuve concluante que le glyphosate augmente la sensibilité des cultures GR aux maladies. Une autre étude (
26 ) remet en question cette évaluation. Plusieurs études ont observé que les betteraves GR et le soja présentent une sensibilité accrue aux agents pathogènes lorsque le glyphosate est appliqué aux doses recommandées (
26 – 28 ). Une étude n’a révélé aucun effet du glyphosate sur l’induction de maladies chez les betteraves GR jusqu’à ce que les taux d’application normaux au champ soient dépassés d’un ordre de grandeur (
29 ). Cependant, d’autres études sur les cultures GR n’ont trouvé aucune influence du glyphosate sur la maladie (
30 ) et même quelques cas d’activité fongicide du glyphosate contre certains agents pathogènes des plantes, en particulier les rouilles (examiné par Duke [
31 ]).Deux études clés ont soutenu l’hypothèse de l’enrichissement en agents pathogènes en glyphosate, révélant sur de longues périodes d’étude que le glyphosate augmente de manière répétée le taux de colonisation des cultures par
Fusarium spp. (présumées être des souches pathogènes) tout en diminuant l’abondance de bactéries fluorescentes
Pseudomonas (considérées comme des organismes bénéfiques putatifs) dans le sol (
28 ). Ces études sont souvent citées comme preuve concluante que l’utilisation à long terme du glyphosate augmente la charge pathogène et diminue l’abondance des bactéries favorisant la croissance dans les sols. Les deux études ont appliqué une méthodologie basée sur la culture pour quantifier ces groupes microbiens, avec une analyse moléculaire de la région d’espacement transcrite interne (ITS) ribosomale pour les champignons. Les études utilisant une méthodologie sans culture pour caractériser les communautés microbiennes n’ont pas réussi à détecter des effets substantiels du glyphosate sur l’abondance des agents pathogènes (
33 ). Les systèmes agricoles, les facteurs pédologiques, les variétés de cultures et l’historique d’utilisation du glyphosate peuvent tous avoir un impact sur le comportement du glyphosate et son interaction avec les microbiomes des cultures et du sol (
34 ) et doivent être inclus dans la conception expérimentale.Nous avons mené deux études à l’échelle du terrain pour déterminer les effets du glyphosate sur le microbiome du sol et la santé des plantes pour les variétés de maïs et de soja GR, testant l’hypothèse selon laquelle le glyphosate modifie la composition du microbiome du sol selon différents types de sol, cultures, moments d’échantillonnage, et les systèmes agricoles. De plus, nous avons testé l’hypothèse selon laquelle
Fusarium sp. l’abondance des séquences ou les nombres cultivables augmenteraient en raison du traitement au glyphosate. Notre étude comprenait six systèmes agricoles étudiés sur deux ans, représentant diverses pratiques agricoles mises en œuvre dans des fermes en activité. Notre étude a ciblé à la fois les microbiomes naïfs du sol qui n’avaient pas été exposés au glyphosate et ceux exposés au glyphosate chaque année. Le séquençage à haut débit a été utilisé pour générer des profils génétiques d’ARNr 16S bactériens et archéens et des profils ITS ribosomiques nucléaires fongiques.
RÉSULTATS
Résumé de la diversité fongique et procaryote sur tous les sites.
À partir de l’analyse de séquençage, un total de 68 964 variantes uniques de séquence d’amplicons fongiques et 72 454 procaryotes uniques (ASV) ont été identifiées dans tous les échantillons. Beltsville, MD, et Stoneville, MS, partageaient 13 964 taxons procaryotes et 5 740 taxons fongiques. Stoneville présentait respectivement 62 985 et 29 780 ASV procaryotes et fongiques. Beltsville présentait respectivement 41 538 et 44 924 ASV procaryotes et fongiques uniques. La diversité fongique était plus élevée à Beltsville qu’à Stoneville, à l’exception des mesures de diversité de Shannon et Simpson pour Org3 (voir la description des rotations sur le terrain dans Matériels et méthodes) (
Fig. 1A ). À l’inverse, la diversité procaryote était plus grande à Stoneville qu’à Beltsville dans toutes les mesures (
FIG 1 Résumé de la diversité des espèces et des structures des communautés bactériennes et fongiques pour tous les échantillons de sol de Beltsville, MD et Stoneville, MS ( n = 768). (A) Estimations de la diversité des espèces pour les champignons dans tous les systèmes agricoles. (B) Diversité des espèces pour les procaryotes dans tous les systèmes agricoles. (C) Analyse de correspondance sans tendance (DCA) de la dissimilarité de Bray-Curtis pour les communautés fongiques de tous les échantillons après transformation de l’abondance relative des comptes totaux. (D) DCA de la dissimilarité de Bray-Curtis pour les communautés procaryotes de tous les échantillons après transformation de l’abondance relative des décomptes totaux.
L’analyse des correspondances sans tendance a montré que les communautés de Beltsville et de Stoneville étaient distinctes (
D ). L’analyse de variance multivariée permutationnelle (PERMANOVA) de l’abondance relative des champignons et des procaryotes a révélé que l’emplacement était le facteur le plus significatif expliquant les différences dans les communautés fongiques et procaryotes du sol (
P = 0,001 dans les deux cas ;
R 2 fongique = 0,19,
R 2 procaryote = 0,16 ; voir le tableau S1 dans le matériel supplémentaire). Les différences entre les communautés microbiennes de Stoneville et de Beltsville étaient dues à des différences dans les facteurs édaphiques. Les caractéristiques chimiques du sol différaient entre les deux emplacements (analyse de discrimination canonique,
P < 0,001,
R 2 = 0,99) et entre les systèmes agricoles (analyse de discrimination canonique,
P < 0,001,
R 2 = 0,99). Le sol de Stoneville avait un pH et des cations As et Sr significativement plus élevés (analyse de variance [ANOVA],
P < 0,001), tandis que le sol de Beltsville contenait significativement plus de P, Pb, S, Fe et de matière organique (MO) (ANOVA ,
P < 0,001). Les ordinations d’échelle multidimensionnelle non métrique (NMDS) de la dissimilarité de Bray-Curtis pour la chimie du sol entre les emplacements et les systèmes agricoles sont présentées à la
Fig . Pour augmenter la puissance de détection des effets locaux du traitement au glyphosate, nous avons analysé les cultures (maïs par rapport au soja) séparément au sein de chaque emplacement (Beltsville par rapport à Stoneville).FIGURE 2
FIG 2 Ordre d’échelle multidimensionnel non métrique de la dissimilarité de Bray-Curtis pour la chimie du sol des sites examinés dans cette étude. Les flèches indiquent les vecteurs des différentes composantes de la composition du sol.
Structure de la communauté fongique et réponse au glyphosate.
Le système agricole était le principal facteur de structure de la communauté fongique, quelle que soit la culture (
R 2 = 0,23). L’année d’échantillonnage était également significative mais expliquait moins de variance que le système agricole de Beltsville (maïs,
P = 0,001,
R 2 = 0,046 ; soja,
P = 0,001,
R 2 = 0,043) et de Stoneville (maïs,
P = 0,001 ). ,
R 2 = 0,051 ; soja,
P = 0,001,
R 2 = 0,052). L’identité taxonomique de la diversité fongique est résumée au niveau de l’ordre sur
la figure 4 (voir également les figures S1 et S5 dans le matériel supplémentaire). Les différences entre les systèmes sont réparties le long de l’axe 1 des graphiques d’analyse des correspondances canoniques (ACC), et les différences liées à l’année sont reflétées dans la répartition le long de l’axe 2 sur
la figure 3 . Aucune interaction significative n’a été notée entre la date d’échantillonnage et le traitement au glyphosate (
P = 0,488 et 0,296 pour le maïs et le soja, respectivement). Les partitions de la rhizosphère (proches et lointaines) n’étaient pas non plus significativement différentes (tableau S1) pour n’importe quelle culture ou emplacement. Les tests de rapport de vraisemblance de l’abondance des taxons dans DESeq2 n’ont également révélé aucune augmentation significative du pouvoir explicatif d’un modèle contenant l’interaction date d’échantillonnage-traitement au glyphosate pour tout taxon, quelle que soit la culture ou le système agricole (Tableaux S4 et S5).FIGURE 3
FIG 3 Analyses canoniques de correspondance (ACC) des communautés fongiques du sol à Beltsville, MD, et Stoneville, MS, réparties par culture. Les données ont été stabilisées en variance avec une transformation binomiale négative dans DESeq2. (A) Parcelles de maïs de Beltsville. (B) Parcelles de soja de Beltsville. (C) Parcelles de maïs de Stoneville. (D) Parcelles de soja Stoneville.
FIGURE 4
FIG. 4 Abondances proportionnelles des communautés fongiques du sol à Beltsville, MD et Stoneville, MS, réparties par culture à partir des données de la Fig. 3 . Taxons dont l’abondance est inférieure à 1 % exclus. Des parcelles avec tous les taxons inclus peuvent être trouvées sur la figure S1 dans le matériel supplémentaire et 5. (A) Parcelles de maïs de Beltsville. (B) Parcelles de soja de Beltsville. (C) Parcelles de maïs de Stoneville. (D) Parcelles de soja Stoneville.
Structure de la communauté procaryote et réponse au glyphosate.
Le système agricole était également significatif pour la structure de la communauté procaryote (
R 2 = 0,16). Le terme annuel expliquait une variance plus faible que le système agricole de Beltsville (maïs,
P = 0,001,
R 2 = 0,096 ; soja,
P = 0,001,
R 2 = 0,086) et de Stoneville (maïs,
P = 0,001,
R 2 = 0,051 ; soja,
P = 0,001,
R 2 = 0,069). Les différences entre les systèmes sont réparties le long de l’axe 1 des parcelles CCA, et les différences liées à l’année sont reflétées dans la répartition le long de l’axe 2 de
la figure 5 . L’identité taxonomique de la diversité procaryote est résumée au niveau de l’ordre sur
les figures 6 , S6 et S10. L’interaction entre le glyphosate et la date d’échantillonnage n’était significative pour aucune des deux cultures (tableau S1). Les tests de rapport de vraisemblance de l’abondance des taxons dans DESeq2 indiquent que l’interaction date d’échantillonnage-traitement au glyphosate n’a pas augmenté de manière significative le pouvoir explicatif du modèle pour aucun taxon, quel que soit la culture ou le système agricole (Tableaux S6 et S7).FIGURE 5
FIG 5 Analyses de correspondance canonique (ACC) des communautés procaryotes du sol à Beltsville, MD, et Stoneville, MS, réparties par culture. Les données ont été stabilisées en variance avec une transformation binomiale négative dans DESeq2. (A) Parcelles de maïs de Beltsville. (B) Parcelles de soja de Beltsville. (C) Parcelles de maïs de Stoneville. (D) Parcelles de soja Stoneville.
FIGURE 6
FIG. 6 Abondances proportionnelles des communautés procaryotes du sol à Beltsville, MD, et Stoneville, MS, réparties par culture à partir des données de la Fig . Les barres ne totalisent pas 1 car les taxons dont l’abondance est inférieure à 1% sont exclus, ce qui peut provoquer l’apparition de taxons manquants parmi les traitements. Des parcelles avec tous les taxons inclus peuvent être trouvées sur les figures S6 et S10. (A) Parcelles de maïs de Beltsville. (B) Parcelles de soja de Beltsville. (C) Parcelles de maïs de Stoneville. (D) Parcelles de soja Stoneville.
Différences de richesse communautaire entre les échantillons avant et après pulvérisation.
Les tests de classement signés de Wilcoxon ont montré plusieurs cas où la diversité des espèces différait de manière significative entre les dates d’échantillonnage avant et après pulvérisation (
Fig. 7 et tableaux S2 et S3); cependant, des différences ont été observées dans les traitements par pulvérisation et sans pulvérisation pour la plupart des combinaisons de systèmes de culture, ce qui indique qu’il s’agit d’un effet saisonnier et non dû à l’exposition au glyphosate. À Beltsville, la réponse du maïs et du soja différait selon les deux dates. La diversité des procaryotes pour le maïs dans chaque système agricole de Beltsville était significativement différente entre les deux dates. Cette tendance a également été observée, mais dans une moindre mesure, dans les communautés fongiques. La moitié des traitements différaient significativement pour les traitements par pulvérisation et sans pulvérisation. Les communautés fongiques ne différaient pas selon les saisons dans les parcelles de soja de Beltsville, et la diversité des espèces fongiques n’était pas affectée par la date d’échantillonnage du maïs et du soja dans les échantillons de Stoneville.FIGURE 7
FIG 7 Modification de la diversité des données raréfiées de Shannon selon les dates d’échantillonnage dans les traitements sans pulvérisation et par pulvérisation. Les astérisques sur chaque parcelle correspondent aux valeurs P brutes (*) et au taux de fausses découvertes corrigées (**) inférieures à 0,05 d’après le test de rang signé de Wilcoxon des différences entre les dates. Les années sont regroupées bien que représentées graphiquement séparément. Les points rouges représentent la diversité moyenne. (A) Diversité fongique de Shannon dans le maïs. (B) Diversité procaryote de Shannon dans le maïs. (C) Diversité fongique de Shannon dans le soja. (D) Diversité procaryote de Shannon dans le soja.
Quantification des UFC de Fusarium .
Le criblage des endophytes racinaires a nécessité l’analyse de plus de 6 100 segments de racines et a identifié plus de 2 400 colonies fongiques. Un nombre significativement plus élevé d’UFC ont été observés en 2013 qu’en 2014 à l’emplacement de Beltsville (
P < 0,0003), mais aucune différence dans le nombre d’UFC n’a été observée entre les années à l’emplacement de Stoneville. Un total de 384 morphotypes typiques ont été séquencés par amplicon ITS, ce qui a donné les 8 taxons dominants identifiés suivants :
Fusarium ,
Macrophomina ,
Alternaria ,
Cladosporium ,
Penicillium ,
Zygomycota ,
Trichoderma et
Epicoccum . Il n’y a aucune différence significative dans les UFC
de Fusarium observées entre les traitements par pulvérisation et sans pulvérisation de glyphosate pour le maïs ou le soja (ANOVA,
P < 0,07). Bien que la valeur
P soit proche du seuil de signification, la variance autour des moyennes ne montre aucune tendance détectable dans les données (
FIG 8 Abondance des isolats de Fusarium ± écart type. (A) Racines de maïs. (B) Racines de soja. Sites du Mississippi et du Maryland analysés séparément avec ANOVA. Les couleurs suivent celles utilisées sur les figures 3 et 4 .
Rendements de maïs et de soja.
Il n’y avait aucune différence significative dans le rendement du maïs selon les systèmes ou entre les traitements d’application de glyphosate pour 2013 ou 2014 (
Tableau 1 ). Les rendements du maïs et du soja dans cette étude ont déjà été publiés (
36 ). Les rendements du maïs n’étaient pas significativement différents des moyennes du comté du MD pour tous les systèmes, avec une moyenne parmi les systèmes de 9 339 kg ha
−1 . En 2013, une erreur s’est produite lors de l’utilisation de la moissonneuse-batteuse pour petites parcelles et les haricots récoltés à partir de différentes répétitions ont été mélangés, rendant les données inutilisables. En 2014, les rendements du soja étaient similaires aux moyennes du comté avec une moyenne de 2 327 kg ha
−1 . Il n’y avait pas de différence significative de rendement entre les systèmes agricoles et aucun effet du traitement au glyphosate sur le rendement (
Tableau 1 ).TABLEAU 1TABLEAU 1 Rendement du maïs et du soja Maryland pour les parcelles traitées ou non au glyphosate en culture ciselée, sans labour, en rotation biologique de 3 ans ou en rotation biologique de 6 ans a
Année
Recadrer
Philosophie de gestion
Travail du sol primaire
Rendement en traitement au glyphosate (kg ha −1 ) b
Vaporisateur
Pas de pulvérisation
2013
Maïs
Conventionnel
Ciseau jusqu’à
8 848
9 536
Conventionnel
Non jusqu’à
9 141
9 780
Biologique 3 ans
Charrue à versoir
7 634
9 210
Biologique 6 ans
Charrue à versoir
8 510
8 832
Soja
Conventionnel
Ciseau jusqu’à
–
–
Conventionnel
Non jusqu’à
–
–
Biologique 3 ans
Charrue à versoir
–
–
Biologique 6 ans
Charrue à versoir
–
–
2014
Maïs
Conventionnel
Ciseau jusqu’à
8 967
9 798
Conventionnel
Non jusqu’à
11 123
10 757
Biologique 3 ans
Charrue à versoir
9 497
10 225
Biologique 6 ans
Charrue à versoir
8 185
7 627
Soja
Conventionnel
Ciseau jusqu’à
2 160
2 438
Conventionnel
Non jusqu’à
2 016
2 264
Biologique 3 ans
Charrue à versoir
2 907
2 315
Biologique 6 ans
Charrue à versoir
2 733
2 290
unLa comparaison des moyennes a été calculée au sein de chaque système pour le génotype résistant au glyphosate, traité ou non avec le glyphosate.
bAucune différence significative n’a été constatée entre les parcelles traitées ou non au glyphosate au sein de chaque système en 2014. –, en 2013, une erreur de récolte des parcelles a entraîné un mélange de parcelles traitées et non traitées, rendant ainsi les données de rendement inutilisables.
DISCUSSION
Les structures des communautés procaryotes et fongiques entre les systèmes agricoles et entre les dates d’échantillonnage n’étaient pas influencées par l’utilisation du glyphosate. Au lieu de cela, le travail du sol et d’autres différences entre les systèmes agricoles semblent être les principaux facteurs déterminants de la structure du microbiome du sol (
5 ). Par exemple, même si tous les champs de Beltsville étaient sous gestion sans labour jusqu’en 1996, les différences de gestion depuis lors sont des indicateurs importants de la structure actuelle de la communauté microbienne. Les systèmes biologiques ont montré une augmentation des champignons de l’ordre des Pezizales (
B ), probablement une réponse des taxons saprobes à l’ajout de litière de volaille dans ces parcelles. L’histoire de l’utilisation des terres à Stoneville différait considérablement d’un système à l’autre, un système étant soumis à une gestion agricole sans labour avec un historique d’application de glyphosate depuis 15 ans et l’autre étant une monoculture de cogongrass sans historique d’exposition au glyphosate. Les différences entre les communautés fongiques dans les parcelles de Stoneville semblent être dues à des proportions changeantes, avec quelques ordres d’abondance supérieure à 5 % présents dans un système alors qu’ils sont absents dans l’autre (
D ).L’héritage de la gestion agricole a déplacé les communautés procaryotes entre les systèmes du Maryland et du Mississippi. L’histoire de la gestion sans labour semble avoir modifié la structure de la communauté microbienne par rapport au labour conventionnel et aux traitements biologiques. Par exemple, les acidobactéries ont été détectées, bien qu’à des niveaux relatifs faibles, dans les proportions les plus élevées dans les systèmes de culture sans labour ayant au moins 15 ans d’expérience. Les acidobactéries réagissent positivement aux nitrates présents dans le sol et il a été démontré qu’elles produisent l’hormone de croissance des plantes, l’acide indole acétique (IAA), qui peut favoriser la croissance des racines des plantes (
Chloroflexi était l’un des taxons les plus réactifs aux changements dans les systèmes agricoles. Un taxon peu connu, les
Ktedonobacteria , a également montré une réponse différentielle aux traitements au niveau du système. Les membres de ce groupe semblent être sensibles au pH, occupent un large éventail d’environnements et sont génétiquement similaires aux
Chloroflexi (
39 ). Les résultats pour les taxons procaryotes et fongiques étudiés dans cette étude sont cohérents avec les différences de systèmes agricoles observées dans d’autres dimensions des écosystèmes du sol à Beltsville, y compris les communautés de nématodes du sol (
40 ), les concentrations de MO et de P dans le sol, les émissions de gaz à effet de serre et les coûts énergétiques totaux. du système agricole (
41 – 43 ).L’absence d’effets du glyphosate dans des communautés de sol auparavant naïves suggère que les taux d’application typiques de glyphosate ne modifient pas la communauté microbienne globale lors de la résolution des taxons récupérés dans notre étude. La littérature existante suggère que la plupart des communautés microbiennes sont susceptibles d’être perturbées, bien que les préjugés contre la déclaration de l’absence d’effets du traitement pourraient affecter ce point de vue (
44 ). Dans la présente étude, la résilience au glyphosate pourrait être liée à plusieurs facteurs. Certaines espèces procaryotes et fongiques sont connues pour métaboliser le glyphosate, et la présence de ces organismes peut protéger les espèces sensibles (
46 ). Les études rapportant les effets du glyphosate sur les microbes du sol utilisent souvent des concentrations d’herbicide plus élevées que le taux approuvé, ce qui peut submerger le tampon des membres résistants. Le glyphosate est fortement lié aux composants du sol (
23 ), mais on ne sait pas comment cela affecte sa biodisponibilité pour les microbes du sol. Néanmoins, sa demi-vie dans les sols à climat tempéré est en moyenne d’environ 30 jours (
47 ). Des effets dépendants de la concentration du glyphosate sur la respiration microbienne et la biomasse du sol ont été rapportés et sont cohérents avec les rapports sur d’autres produits agrochimiques, ne montrant que des effets transitoires aux taux d’application recommandés (
48 ).Des études en serre avec du blé GR cultivé dans des sols de tout le nord-ouest du Pacifique n’ont révélé que des effets mineurs du glyphosate sur les communautés microbiennes, et l’emplacement déterminé était un facteur majeur de la structure de la communauté microbienne du sol (
33 ). Bien que ces études aient détecté de légers effets du glyphosate sur les communautés microbiennes, le glyphosate a été appliqué à deux fois la dose recommandée, augmentant ainsi la probabilité que la communauté microbienne subisse un effet détectable. Cette différence méthodologique peut expliquer la détection d’un effet sur l’abondance de certains taxons après exposition au glyphosate pour le blé cultivé en serre, alors qu’aucun n’a été détecté dans notre étude. Cela renforce la confiance dans notre conclusion selon laquelle le glyphosate a un effet minimal sur la communauté microbienne lorsqu’il est appliqué au taux recommandé.La diversité des communautés a changé au cours de la saison de croissance, quelle que soit l’application de glyphosate (
Fig. 7 ). Ces résultats sont similaires à ceux de Hart et al. (
49 ) qui ont cultivé du maïs GR et son isoline génétiquement proche avec et sans application de glyphosate pendant une saison au Canada. En utilisant le polymorphisme de longueur des fragments de restriction terminaux (TRFLP) pour comparer les communautés microbiennes, ils ont également montré des changements dans la diversité des communautés microbiennes au fil du temps, mais pas en relation avec le glyphosate.Des travaux antérieurs basés sur la culture ont révélé que l’abondance
de Fusarium augmentait et celle
de Pseudomonas diminuait avec l’utilisation de glyphosate (
Fusarium spp. étaient présumés pathogènes, tandis que
Pseudomonas spp. étaient présumés être des symbiotes. Notre métabarcodage n’a détecté aucun effet du glyphosate sur l’abondance de
Fusarium ou
de Pseudomonas sp. (voir le matériel supplémentaire).Il est important de noter que les gènes d’ARNr ITS et 16S ne parviennent pas à résoudre les classifications au niveau des espèces pour certains groupes (
Metarhizium connues pour être présentes sur le site du Maryland (
53 ) n’étaient pas représentées dans les échantillons de cette étude. Très probablement, des agents pathogènes des plantes ont été omis dans cette étude. Cependant, les espèces pathogènes contribuent à l’abondance relative de leur OTU constitutive et nous n’avons détecté aucun changement dans les abondances relatives de
Fusarium sp.,
Alternaria sp. ou
Macrophomina sp. Les OTU augmentent en raison de l’application de glyphosate (voir le matériel supplémentaire). Bien que
Pseudomonas spp. sont souvent considérés comme intrinsèquement bénéfiques, il existe au moins quelques agents pathogènes confirmés (
51 ) et le type de fonction bénéfique peut différer considérablement selon les souches. Quoi qu’il en soit, comme pour les champignons, aucun
Pseudomonas spp. la prévalence a changé à la suite du traitement au glyphosate.Nous n’avons également constaté aucune réduction du rendement par l’application de glyphosate sur le maïs GR ou le soja GR dans les champs ayant une longue histoire d’utilisation du glyphosate ou sans historique d’utilisation du glyphosate (
Tableau 1 ). Dans une étude similaire portant sur le maïs sucré GR, il y a même eu une légère augmentation du rendement associée à l’application de glyphosate (
54 ). Cela pourrait être dû à l’hormèse, où des doses non phytotoxiques de glyphosate stimulent la croissance des plantes (
55 ). L’absence d’effets sur les rendements concorde avec l’absence d’effets néfastes substantiels sur les microbes de la rhizosphère.Le glyphosate est l’herbicide le plus utilisé dans le monde et les cultures GR sont les cultures transgéniques les plus utilisées (
11 ). Aux États-Unis, plus de 90 % des terres agricoles cultivées en soja, coton, betterave sucrière et maïs sont plantées en cultivars GR (
56 ). En 2014, les cultures GR ont reçu 88 % du glyphosate utilisé dans l’agriculture américaine. L’adoption du soja GR et l’utilisation intensive de glyphosate qui en découle en Argentine et au Brésil ont suivi une tendance similaire à celle des États-Unis (
11 ). Bien que les rendements du maïs et du soja aux États-Unis continuent d’augmenter à peu près aux mêmes rythmes qu’avant l’introduction des cultures GR (
57 ), une quantité importante de littérature suggère que le glyphosate devrait compromettre les cultures GR en modifiant négativement les populations de microbes du sol (voir, par exemple). , références
24 à 26 ). Cependant, bon nombre des études soutenant ce point de vue n’ont pas été menées dans des situations agricoles réalistes. Par exemple, l’une des études citées comme preuve des forts effets du glyphosate sur les microbes associés aux plantes a été réalisée dans une serre sur des plantes cultivées en culture hydroponique (
58 ). Relativement peu d’études ont étudié l’effet du glyphosate sur les communautés microbiennes du sol dans les systèmes agricoles avec ou sans héritage d’application de glyphosate. Les travaux décrits dans le présent article apportent une contribution importante à la détermination de l’effet du glyphosate sur les communautés procaryotes et fongiques du sol, car il s’agit d’une étude bien reproduite (dans le temps et dans l’espace) sur deux sites géographiquement séparés dans des systèmes agricoles réalistes avec des antécédents documentés d’utilisation du glyphosate. Le fait qu’il n’y ait aucun changement dû au glyphosate, associé à une tendance vers une plus grande diversité d’espèces dans les parcelles sans labour, suggère que cette pratique de gestion largement utilisée ne risque pas d’altérer les communautés microbiennes du sol de manière négative.
MATÉRIELS ET MÉTHODES
Conditions de terrain et conception expérimentale.
L’étude a été menée en 2013 et 2014 sur deux sites du Département américain de l’agriculture, du Service de recherche agricole (USDA-ARS), au Laboratoire des systèmes agricoles durables de Beltsville, dans le Maryland, et à l’Unité de recherche sur les systèmes de production végétale à Stoneville, MS (
Tableau 2 ). .TABLEAU 2TABLEAU 2 Description des systèmes agricoles représentés dans les expériences sur le terrain à Beltsville, MD, et Stoneville, MS, en 2013 et 2014a
Emplacement
Philosophie de gestion
Travail du sol primaire
Histoire du glyphosate
Nom du système agricole
Beltsville
Conventionnel
Ciseau jusqu’à
Oui
CT
Conventionnel
Non jusqu’à
Oui
NT
Rotation biologique de 3 ans
Charrue à versoir
Non
Organisation_3
Rotation biologique de 6 ans
Charrue à versoir
Non
Organisation_6
Pierreville
Conventionnel
Non jusqu’à
Oui
NT_15 ans
Conventionnel
Non jusqu’à
Non
NT_Aucun
unChaque système agricole était représenté par du maïs et du soja résistants au glyphosate (GR), et chacun n’était pas traité ou traité au glyphosate à raison de 0,87 kg ha
−1 .Le site de Beltsville est géré dans le cadre d’un site de recherche agroécologique à long terme de l’USDA qui comprend des systèmes agricoles typiques de la région médio-atlantique décrits précédemment (
59 ). Nous avons mené l’étude dans deux systèmes agricoles conventionnels, dont un utilisant une charrue chisel pour le labour primaire (CT) et un autre en gestion sans labour (NT). Ces deux systèmes s’appuient sur des engrais minéraux, des herbicides et d’autres pesticides selon les besoins pour gérer une rotation maïs (
Zea mays )-seigle (
Secale céréales )-soja (
Glycine max )-blé d’hiver (
Triticum aestivum )/soja. De plus, deux systèmes biologiques ont été utilisés sur ce site. Un système biologique est une rotation de 3 ans maïs-seigle-culture de couverture-soja-blé d’hiver/vesce velue (
Vicia villosa ) (Org3). La seconde est une rotation de cultures de 6 ans (Org6) dans laquelle la luzerne (
Medicago sativa ), culture pérenne en place depuis 3 ans, remplace la vesce présente dans Org3. Les systèmes biologiques reposent sur les légumineuses, la litière de volaille et le K
2 SO
4 pour fournir des éléments nutritifs aux cultures conformément aux résultats des analyses de sol et aux réglementations locales. Une charrue à versoir et/ou à burin est utilisée pour le travail du sol primaire, et le contrôle des mauvaises herbes comprenait l’utilisation d’une houe rotative et le travail entre les rangs après la plantation de maïs et de soja dans les systèmes biologiques.À Stoneville, l’expérience est composée de deux systèmes agricoles établis dans deux champs adjacents, l’un avec un héritage d’utilisation du glyphosate depuis 15 ans (NT_15yrs) et l’autre sans historique de glyphosate (NT_none). Quatre répétitions ont été délimitées dans chaque champ pour chaque système agricole. Le champ ayant des antécédents d’utilisation du glyphosate cultivait en rotation du soja GR et du coton (
Gossypium hirsutum ) au cours des 15 dernières années précédant l’expérience. Le champ sans historique de glyphosate avait été entretenu pour des études de biologie des mauvaises herbes dans une monoculture de cogongrass (
Imperata cylindrica ) sans herbicides appliqués pendant 12 ans avant l’expérience. La préparation du champ comprenait l’abattage du cogonggrass avec un travail du sol répété, la plantation de soja et de maïs non-GR pendant une saison avant l’expérience sur le terrain en cours et le fauchage au fléau à maturité. Au cours de l’expérience, chaque champ (NT_15yrs ou NT_none) a été divisé en deux, une moitié étant plantée en maïs et l’autre moitié en phase de soja de l’expérience. L’année suivante, des parties du champ qui avaient été ensemencées en maïs ont été ensemencées en soja et vice versa.L’expérience a été menée pendant les phases de rotation des cultures de maïs et de soja aux deux endroits. À chaque emplacement de chaque système agricole et combinaison de cultures, les traitements au glyphosate suivants ont été établis : un cultivar GR sans glyphosate appliqué, et le même cultivar GR avec du glyphosate appliqué à raison de 0,87 kg ha
−1 deux fois 4 semaines après la plantation. Il y a eu deux événements d’échantillonnage pour chaque unité expérimentale. Des échantillons de sol et de racines ont été prélevés au moment du « prétraitement », soit au stade de croissance V4. Le lendemain, du glyphosate a été appliqué sur les parcelles prévues pour recevoir du glyphosate. Environ 20 jours plus tard, un échantillon « postspray » a été prélevé dans chaque unité expérimentale. Les unités expérimentales à tous les emplacements étaient constituées de quatre rangées de 4,6 m de largeur et 6,1 m de longueur. Le cultivar de soja USG Allen (GR) a été semé à 526 400 graines ha
-1 , et le cultivar de maïs DKC 65-17 RR2 (GR) a été semé à 67 600 graines ha
-1 . À Beltsville, les parcelles de maïs ou de soja constituent chacune une phase de la rotation des parcelles principales qui est un système agricole (NT, CT, Org3 ou Org6) ; ainsi, chaque phase de la rotation est considérée comme une parcelle divisée de la parcelle principale. Aux deux endroits, quatre répétitions de chaque facteur et niveau ont été établies. Toutes les parcelles ont été débarrassées des mauvaises herbes grâce à un binage manuel selon les besoins.En octobre de chaque année, sur les deux sites, le maïs a été récolté avec une moissonneuse-batteuse pour petites parcelles Almaco (Nevada, IA) ; le rendement en grains a été calculé à 15,5 % d’humidité pour les deux rangées centrales des parcelles de 6,1 m. Au cours des deux années, sur le site de Stoneville, le soja a été récolté avec une moissonneuse-batteuse Almaco pour petites parcelles. À Beltsville, en 2013, le soja a été récolté avec une moissonneuse-batteuse Almaco pour petites parcelles, et en 2014, le soja a été récolté à la main et battu sur 3,05 m des deux rangées centrales. Les poids secs ont été calculés à 13,5 % d’humidité.
Caractéristiques de base du sol.
Les sols de Beltsville sont des ultisols limoneux limoneux des plaines côtières constitués principalement d’unités cartographiques de sol Christiana, Keyport, Matapeake et Mattapex. Les sols de Stoneville étaient un loam limoneux typique de l’Alfisol dominé par les unités cartographiques des sols de Dundee. Lors de la plantation, des échantillons de sol provenant des 15 premiers cm de profondeur ont été collectés dans chaque parcelle en combinant le sol de six carottes ou plus (7,5 cm de diamètre) échantillonnées de manière semi-aléatoire dans une parcelle donnée. Les échantillons ont été séchés à l’air et tamisés à 2 mm. Les carottes ont été collectées sur une ligne diagonale entre le deuxième et le troisième rang de culture, à 1 m de chaque extrémité d’une parcelle donnée. Des échantillons de sol ont été analysés par le laboratoire des services d’analyse agricole de l’université d’État de Pennsylvanie pour déterminer le pH, la teneur en matière organique (MO), la capacité d’échange cationique (CEC) et P, K, Mg, Ca, S, B, Zn, Mn, Fe, Teneur en Cu, As, Al, Ba, Cd, Co, Cr, Ni, Pb, Se et Sr. Le pH a été déterminé dans une dilution d’eau 1: 1, la MO a été déterminée par perte de masse lors de la combustion et la CEC a été déterminée en utilisant les méthodes de Ross et Ketterings (
60 ). Des extractions Mehlich 3 ont été réalisées pour obtenir du Ca, du Mg et du K du sol ; tous les autres métaux sont exprimés en éléments sorbés totaux selon la méthode EPA 3050 (
Échantillonnage du sol et des racines de la rhizosphère.
Aux stades de croissance V3 à V4 (4 à 6 semaines après la plantation) et 1 jour avant l’application de glyphosate (pré-pulvérisation), six plantes et le sol associé à leurs racines ont été excavés de chaque parcelle en enlevant les monolithes de sol d’un diamètre de 30 cm ( tige de la culture au centre) et à 15 cm de profondeur à l’aide de pelles de précision stérilisées en surface. Les monolithes ont été placés sur un tamis, et la terre autour de la motte a été doucement retirée par agitation et passée à travers un tamis de 2 mm, appelé ici « terre de rhizosphère lointaine ». Le sol adhérant aux racines après cette procédure a été brossé sur un tamis de 2 mm à l’aide d’une brosse en poils de chameau, appelé ici « sol proche de la rhizosphère ». Les racines ont été soigneusement brossées sans compromettre l’intégrité de la surface des racines. Des échantillons de rhizosphère provenant des six plantes ont été regroupés et 5 g ajoutés à un tube Falcon de 15 ml contenant 10 ml de solution de conservation d’acide nucléique Mo Bio LifeGuard. Le contenu des tubes a été mélangé et congelé à –80°C. Les plantes ont été placées à 4°C jusqu’à la poursuite du traitement.
Identification des endophytes à partir des racines.
Des sections de racines de deux centimètres ont été coupées au hasard 16 fois dans chacun des six systèmes racinaires frais pour chaque traitement. Le poids humide total des 16 sections a été enregistré. Les coupes ont été stérilisées en surface pendant 2 minutes dans de l’hypochlorite de sodium à 1,25 %, suivies de trois rinçages à l’eau distillée stérile. Les coupes ont été séchées sur du papier absorbant stérile et huit coupes de racines ont été placées sur une plaque contenant le milieu de Komada (
62 ). Les racines plaquées ont été incubées à la lumière ambiante et à température ambiante jusqu’à l’émergence des colonies. Les mycéliums fongiques et les spores des colonies émergentes ont été échantillonnés et examinés au microscope Nikon E60 et identifiés au niveau du genre ou à des groupes morphologiques plus larges, sur la base de caractéristiques taxonomiques. Des colonies de morphologie typique ont été étalées sur un milieu minimal pour induire la sporulation en vue d’une identification plus approfondie. Des tests PCR pour les ITS, suivis d’un clonage et d’un séquençage, ont été réalisés sur plus de 384 colonies de morphologie typique pour valider l’identification microscopique. Les méthodes suivaient celles décrites dans la référence
63 . La qualité des séquences a été vérifiée et alignée à l’aide du logiciel de la suite DNAStar (DNAStar, Madison, WI, USA) et identifiée à l’aide de l’outil de recherche d’alignement local de base et de la banque de données nucléotidiques GenBank du National Center for Biotechnology Information à Bethesda, MD (
Préparation de bibliothèques de séquençage Illumina à partir de sols de rhizosphère.
La rhizosphère et les sols en vrac conservés dans LifeGuard à –80 °C ont été décongelés et 800 µl de chaque boue ont été traités à l’aide d’un kit d’isolement d’ADN de sol à 96 puits PowerSoil-htp (Mo Bio Laboratories, Inc., Solana Beach, Californie), selon aux recommandations du fabricant. L’ADN a été quantifié et sa qualité vérifiée à l’aide d’un spectrophotomètre NanoDrop 2000 (Thermo Fisher Scientific, Pittsburgh, PA). Des amplicons du gène de l’ARNr bactérien 16S ont été générés avec les amorces f515 et r806 (
64 ). Des amplicons fongiques ITS ont été générés avec les amorces ITS1 et ITS2 (
65 ). Le séquençage du gène de l’ARNr 16S et du métacode à barres ITS a été effectué conformément au manuel de préparation de la bibliothèque de protocoles Illumina (numéro de pièce 1504423 rév. B ; Illumina, Inc.). Cinq microlitres de produit d’amplicon adaptateur nettoyé pour chaque échantillon ont été utilisés pour la PCR d’indexation à l’aide du kit d’indexation Nextera XT (référence FC-131-1002 ; manuel de préparation de la bibliothèque de séquençage métagénomique 16S, référence 1504423 rév. B ; Illumina, Inc.) . Les produits de PCR index ont été nettoyés conformément au protocole Illumina (manuel de préparation de la bibliothèque de séquençage métagénomique 16S, référence 1504423 rév. B ; Illumina, Inc.), et des aliquotes de 2 µl par échantillon de chaque plaque PCR à 96 puits ont été regroupées pour le résultat final. Bibliothèque Illumina. Pour l’analyse, 100 µl de solutions à 10 nM de chaque pool de bibliothèques ont été congelés et expédiés sur de la neige carbonique pour analyse sur un système Illumina MiSeq au Centre de recherche sur le génome et de bioinformatique (CGRB), Oregon State University, Corvallis, OR. Pour Beltsville, un total de 512 échantillons ont été séquencés. Pour Stoneville, 256 échantillons ont été séquencés.
Bioinformatique et analyse statistique.
(i) Filtrage et découpage des séquences. Les lectures ont été renvoyées par le CGRB après un contrôle de qualité initial avec les flux de travail Illumina standard, y compris le filtrage de qualité et le découpage de l’adaptateur. Les scripts utilisés dans les étapes suivantes peuvent être trouvés sur
https://github.com/rmkepler/FSP_script_repository . Avant de joindre les extrémités appariées et l’attribution de la taxonomie, les amorces directes et inverses ont été supprimées et la qualité des séquences réduite (-q 22) à l’extrémité 3 ‘à l’aide de Cutadapt (version 1.8.3) (
66 ). Les lectures dépourvues de séquences d’amorces ou inférieures à 75 pb avant le découpage ont été rejetées.
(ii) Affectation d’assemblage et de taxonomie. Le package R Dada2 (
67 ) a été utilisé pour l’assemblage par paires et l’affectation de taxonomie. La commande « filterandtrim » a été utilisée pour supprimer les séquences avec un taux d’erreur attendu > 2 et toutes les séquences contenant des valeurs « N » (bases illisibles). Les taux d’erreur ont été estimés pour les lectures directes et inverses. Les lectures filtrées ont ensuite été dérépliquées avec la commande « derepFastq ». Les séquences dérépliquées ont été débruitées avec la commande « dada », puis les extrémités appariées ont été fusionnées. Les séquences chimériques ont été supprimées avec la commande « removeBimeraDenovo ». La taxonomie a été attribuée à la table de séquences sans chimère avec l’implémentation dada2 du classificateur RDP (
(iii) Analyse communautaire. Nous avons transformé les données sur le gène de l’ARNr 16S et le nombre de communautés ITS en abondances relatives, puis avons calculé la dissimilarité de Bray-Curtis. L’analyse des correspondances sans tendance (DCA) a été appliquée à la matrice de dissimilarité de Bray-Curtis à l’aide du package VEGAN v. 2.4 (
72 ) pour les codes-barres fongiques et procaryotes. PERMANOVA a été utilisé pour évaluer l’importance des facteurs liés à la culture et à l’emplacement.Après un sous-ensemble par culture et par emplacement, la richesse et la régularité ont été estimées à partir d’ensembles de données raréfiées des décomptes de séquences brutes à l’aide de VEGAN. DESeq2 v. 1.18.1 (
73 ) a été utilisé pour produire des ensembles de données à variance stabilisée (
74 ) à partir de décomptes non raréfiés. Les ordinations ont été réalisées à l’aide de l’analyse des correspondances canoniques (ACC) et d’un modèle de la forme « ∼ système + année » avec VEGAN. Nous avons utilisé PERMANOVA pour déterminer l’importance des principaux effets et interactions entre les facteurs suivants : système agricole, zone de sol, traitement au glyphosate, date d’échantillonnage et année. Le facteur du système agricole comportait 4 catégories pour Beltsville (CT, NT, Org3 et Org6) et 2 pour Stoneville (NT_none et NT_15yr). Tous les autres facteurs avaient deux catégories aux deux endroits, comme suit : zone de sol (en vrac et rhizosphère), année (2013 et 2014), traitement au glyphosate (pulvérisation et non pulvérisation) et date d’échantillonnage (application de glyphosate avant pulvérisation et application de glyphosate après pulvérisation). Un modèle à mesures répétées basé sur l’identification des parcelles (ID) a été utilisé.L’effet du traitement au glyphosate sur les communautés microbiennes a été testé avec le test de rang signé de Wilcoxon sur les différences entre les dates, tel que mis en œuvre dans le plug-in longitudinal pour Qiime2 (
75 ). Le test a été appliqué séparément pour trois mesures de diversité pour les données à variance stabilisée, déterminées avec VEGAN, observé, l’indice de Shannon et l’indice de Simpson.
(iv) Taxons différentiellement abondants. Des tests pour des taxons différentiellement abondants en réponse au traitement au glyphosate ont été réalisés dans DESeq2 en utilisant des tests de rapport de vraisemblance après avoir sous-divisé les données fongiques et procaryotes par emplacement, culture et système agricole. Le test a comparé un modèle complet comprenant le groupe, les termes de date d’échantillonnage et un terme d’interaction, où le groupe est défini comme la combinaison du système agricole et du traitement au glyphosate (par exemple, Org3_spray) et la date d’échantillonnage correspond aux événements d’échantillonnage avant et après pulvérisation. Le modèle complet a été comparé à un modèle réduit dépourvu du terme d’interaction. Ainsi, les taxons présentant des valeurs
P significatives indiquent que la date d’échantillonnage et l’application de glyphosate ont interagi pour être d’importants prédicteurs de leur abondance. Cela a été testé pour chaque taxon fongique et procaryote identifié. Des ensembles de données avec des décomptes non transformés ont été utilisés comme données de départ, dont la variance a ensuite été stabilisée pendant les tests.Les différences dans la chimie du sol ont été évaluées par analyse discriminante canonique à l’aide du package candisc v. 0.8.0 dans R. Les différences dans la chimie du sol ont été visualisées avec une mise à l’échelle multidimensionnelle non métrique de la dissimilarité de Bray-Curtis et des vecteurs tracés pour les différents constituants chimiques à l’aide du métaMDS et envfit fonctions de VEGAN, respectivement.
Disponibilité des données.
Les données sont accessibles sous le numéro NCBI BioProject
Chris Rasmann et Gwen Bagley ont contribué de manière substantielle à la gestion sur le terrain et à l’échantillonnage des parcelles de Beltsville. Le regretté John Lydon a contribué à la conception expérimentale.Nous ne déclarons aucun conflit d’intérêts.
note de bas de page
[Cet article a été publié le 18 février 2020, avec le suffixe du nom de Martin M. Williams II affiché à tort comme « Jr. » dans la signature et les références 35, 36 et 54. Cela a été corrigé dans la version de l’article publiée le 18 septembre 2020. De plus, le prénom de Jeffrey S. Buyer était affiché à tort comme « Jeffery », et cela a été corrigé. dans la version de l’article publiée le 14 octobre 2020.]
ASM ne détient pas les droits d’auteur sur le matériel supplémentaire qui peut être lié à un article ou accessible via celui-ci. Les auteurs ont accordé à ASM une licence mondiale non exclusive pour publier les fichiers de matériel supplémentaire. Veuillez contacter directement l’auteur correspondant pour une réutilisation.
LES RÉFÉRENCES
1.Cardoso IM, Kuyper TW. 2006. Mycorhizes et fertilité des sols tropicaux.
Agric Ecosyst Environ 116 : 72-84.
Google Scholar2.Muñoz N, Qi X, Li MW, Xie M, Gao Y, Cheung MY, Wong FL, Lam HM. 2016. L’amélioration de la capacité de fixation de l’azote pourrait faire partie du processus de domestication du soja.
Hérédité (Edinb) 117 : 84-93.
Google Scholar3.Dodd IC, Zinovkina NY, Safronova VI, Belimov AA. 2010. Médiation rhizobactérienne du statut hormonal des plantes.
Ann Appl Biol 157 : 361-379.
Google Scholar4.Kepler RM, Maul JE, Rehner SA. 2017. Gestion du microbiome végétal pour les champignons de biocontrôle : exemples d’Hypocreales.
Curr Opin Microbiol 37 : 48-53.
Google Scholar5.Bakker MG, Manter DK, Sheflin AM, Weir TL, Vivanco JM. 2012. Exploiter le microbiome de la rhizosphère grâce à la sélection végétale et à la gestion agricole.
Sol végétal 360 : 1-13.
Google Scholar6.Mueller UG, Sachs JL. 2015. Ingénierie des microbiomes pour améliorer la santé des plantes et des animaux.
Tendances Microbiol 23 : 606-617.
Google Scholar7.Cohen, J.E.. 2003. Population humaine : le prochain demi-siècle.
Sciences 302 : 1172-1175.
Google Scholar8.Hartman K, van der Heijden MGA, Wittwer RA, Banerjee S, Walser JC, Schlaeppi K. 2018. Les pratiques agricoles manipulent les modèles d’abondance des membres du microbiome des racines et du sol, ouvrant la voie à une agriculture intelligente.
Microbiome 6:14.
Google Scholar9.Zinger L, Bonin A, Alsos IG, Bálint M, Bik H, Boyer F, Chariton AA, Creer S, Coissac E, Deagle BE, Barba MD, Dickie IA, Dumbrell AJ, Ficetola GF, Fierer N, Fumagalli L, Gilbert MTP , Jarman S, Jumpponen A, Kauserud H, Orlando L, Pansu J, Pawlowski J, Tedersoo L, Thomsen PF, Willerslev E, Taberlet P. 2019. Métabarcoding ADN : nécessité de modèles expérimentaux robustes pour tirer des conclusions écologiques solides.
Mol Ecol 28 : 1857-1862.
Google Scholardix.Busby PE, Soman C, Wagner MR, Friesen ML, Kremer J, Bennett A, Morsy M, Eisen JA, Leach JE, Dangl JL. 2017. Priorités de recherche pour exploiter les microbiomes végétaux dans une agriculture durable.
PLoS Biol 15 :e2001793.
Google Scholar11.Benbrook CM. 2016. Tendances de l’utilisation de l’herbicide glyphosate aux États-Unis et dans le monde.
Environ Sci Eur 28:3.
Google Scholar12.Duc SO, Powles SB. 2008. Glyphosate : un herbicide unique en son genre.
Google Scholar13.Wardle DA. 1995. Impacts des perturbations sur les réseaux trophiques de détritus dans les agro-écosystèmes dus à des pratiques contrastées de travail du sol et de gestion des mauvaises herbes.
Adv Ecol Rés 26 : 105-185.
Google Scholar14.Frey SD, Elliott ET, Paustian K. 1999. Abondance et biomasse bactériennes et fongiques dans les agroécosystèmes conventionnels et sans labour le long de deux gradients climatiques.
Sol Biol Biochem 31 : 573-585.
Google Scholar15.Steinrücken HC, Amrhein N. 1980. L’herbicide glyphosate est un puissant inhibiteur de l’acide 5-énolpyruvylshikimique-3-phosphate synthase.
Biochem Biophys Res Commun. 94 : 1207-1212.
Google Scholar16.Dill GM, Sammons Rd, Feng PCC, Kohn F, Kretzmer K, Mehrsheikh A, Bleeke M, Honegger JL, Farmer D, Wright D, Haupfear EA. 2010. Glyphosate : découverte, développement, applications et propriétés, p 1–33.
Dans Nandula VK (éd.),
Résistance au glyphosate dans les cultures et les mauvaises herbes . John Wiley & Sons, Ltd., New York, État de New York.
Google Scholar17.Borggaard OK, Gimsing AL. 2008. Devenir du glyphosate dans le sol et possibilité de lessivage dans les eaux souterraines et de surface : un examen.
Gestion antiparasitaire Sci 64 : 441-456.
Google Scholar18.Johal GS, Rahé JE. 1984. Effet des champignons phytopathogènes du sol sur l’action herbicide du glyphosate sur les semis de haricots.
Phytopathologie 74 : 950-955.
Google Scholar19.Hammerschmidt R. 2018. Comment le glyphosate affecte le développement des maladies des plantes : c’est plus qu’une susceptibilité accrue.
Dans Kearney PC, Kaufman DD (éd.), Herbicides – chimie, dégradation et mode d’action. Marcel Dekker, Inc., New York, New York.
Google Scholar21.Coupland D, Caseley JC. 1979. Présence d’une activité 14c dans les exsudats racinaires et le liquide de guttation d’
Agropyron repens traités avec du glyphosate marqué au
14C .
Nouveau Phytole 83 : 17-22.
Google Scholar22.Laitinen P, Rämö S, Siimes K. 2007. Translocation du glyphosate des plantes vers le sol – cela constitue-t-il une proportion significative des résidus dans le sol ?
Sol végétal 300 : 51-60.
Google Scholar23.Torstensson L. 1985. Comportement du glyphosate dans les sols et sa dégradation.
Google Scholar24.Kremer RJ, signifie NE. 2009. Interactions du glyphosate et des cultures résistantes au glyphosate avec les micro-organismes de la rhizosphère.
Eur J Agron 31 : 153-161.
Google Scholar25.Van Bruggen AHC, He MM, Shin K, Mai V, Jeong KC, Finckh MR, Morris JG. 2018. Effets environnementaux et sanitaires de l’herbicide glyphosate.
Sci Total Environ 616-617 : 255-268.
Google Scholar26.Martinez DA, Loening UE, Graham MC. 2018. Impacts des herbicides à base de glyphosate sur la résistance aux maladies et la santé des cultures : une revue.
Environ Sci Eur 30:2.
Google Scholar27.Larson RL, Hill AL, Fenwick A, Kniss AR, Hanson LE, Miller SD. 2006. Influence du glyphosate sur la pourriture des racines
à Rhizoctonia et
Fusarium dans la betterave sucrière.
Pest Management Sci 62 : 1182-1192.
Google Scholar28.Zobiole LHS, Kremer RJ, Oliveira RS, Constantin J. 2011. Le glyphosate affecte les micro-organismes des rhizosphères du soja résistant au glyphosate.
J Appl Microbiol 110 : 118-127.
Google Scholar29.Barnett KA, Sprague CL, Kirk WW, Hanson LE. 2012. Influence du glyphosate sur la pourriture du collet et des racines à Rhizoctonia (
Rhizoctonia solani ) chez la betterave sucrière résistante au glyphosate.
Mauvaises herbes Sci 60 : 113-120.
Google Scholar30.Kandel YR, Bradley CA, Wise KA, Chilvers MI, Tenuta AU, Davis VM, Esker PD, Smith DL, Licht MA, Mueller DS. 2015. Effet de l’application de glyphosate sur le syndrome de mort subite du soja résistant au glyphosate dans des conditions de terrain.
Plante Dis 99 : 347-354.
Google Scholar31.Duc SO. 2018. Interaction des pesticides chimiques et des ingrédients de leur formulation avec les microbes associés aux plantes et aux ravageurs des plantes.
J Agric Food Chem 66 : 7553–7561.
Google Scholar32.Schlatter DC, Yin C, Burke I, Hulbert S, Paulitz T. 2017. L’emplacement, la proximité des racines et l’historique d’utilisation du glyphosate modulent les effets du glyphosate sur les réseaux de communautés fongiques du blé.
Microbe Ecol 76 : 240-257.
Google Scholar33.Schlatter DC, Yin C, Hulbert S, Burke I, Paulitz T. 2017. Les impacts de l’utilisation répétée du glyphosate sur les bactéries associées au blé sont faibles et dépendent de l’historique d’utilisation du glyphosate.
Application Environ Microbiol 83:e01354-17.
Google Scholar34.Nguyen DB, Rose MT, Rose TJ, Morris SG, van Zwieten L. 2016. Impact du glyphosate sur la biomasse microbienne du sol et la respiration : une méta-analyse.
Sol Biol Biochem 92 : 50-57.
Google Scholar35.Reddy KN, Cizdziel JV, Williams MM, Jr, Maul JE, Rimando AM, Duke SO. 2018. La technologie de résistance au glyphosate a un effet minime, voire nul, sur la teneur en minéraux et le rendement du maïs.
J Agric Food Chem 66 : 10139-10146.
Google Scholar36.Duke SO, Rimando AM, Reddy KN, Cizdziel JV, Bellaloui N, Shaw DR, Williams MM, Jr, Maul JE. 2018. Absence d’effets des transgènes et du glyphosate sur le rendement et la teneur en minéraux et en acides aminés du soja résistant au glyphosate.
Pest Management Sci 74 : 1166-1173.
Google Scholar37.Kielak AM, Barreto CC, Kowalchuk GA, van Veen JA, Kuramae EE. 2016. L’écologie des acidobactéries : aller au-delà des gènes et des génomes.
Microbiol avant 7:744.
Google Scholar38.Ishaq SL, Johnson SP, Miller ZJ, Lehnhoff EA, Olivo S, Yeoman CJ, Menalled FD. 2017. Impact des systèmes de culture, de l’inoculum du sol et de l’identité des espèces végétales sur la structure de la communauté bactérienne du sol.
Microbe Ecol 73 : 417-434.
Google Scholar39.Yabe S, Sakai Y, Abe K, Yokota A. 2017. Diversité des
Ktedonobacteria avec une morphologie de type actinomycètes dans les environnements terrestres.
Microbes Environ 32 : 61-70.
Google Scholar40.Treonis AM, Unangst SK, Kepler RM, Buyer JS, Cavigelli MA, Mirsky SB, Maul JE. 2018. Caractérisation des communautés de nématodes du sol dans trois systèmes de culture grâce à des approches morphologiques et de métabarcoding ADN.
Sci Rep 8 : 2004.
Google Scholar41.Spargo JT, Cavigelli MA, Mirsky SB, Maul JE, Meisinger JJ. 2011. Azote minéralisable du sol et matière organique labile du sol dans divers systèmes de culture à long terme.
Nutr Cycl Agroecosyst 90 : 253-266.
Google Scholar42.Hoffman E, Cavigelli MA, Camargo G, Ryan M, Ackroyd VJ, Richard TL, Mirsky S. 2018. Utilisation d’énergie et émissions de gaz à effet de serre dans la production de céréales biologiques et conventionnelles : prise en compte des apports de nutriments.
Système agricole 162 : 89-96.
Google Scholar43.Cavigelli MA, Teasdale JR, Conklin AE. 2008. Performance agronomique à long terme des grandes cultures biologiques et conventionnelles dans la région médio-atlantique.
Agron J 100 : 785-794.
Google Scholar44.Shade A, Peter H, Allison SD, Baho D, Berga M, Buergmann H, Huber DH, Langenheder S, Lennon JT, Martiny JB, Matulich KL, Schmidt TM, Handelsman J. 2012. Fondamentaux de la résistance et de la résilience des communautés microbiennes.
Microbiol avant 3:417.
Google Scholar45.Shinabarger DL, Braymer HD. 1986. Catabolisme du glyphosate par
Pseudomonas sp. souche PG2982.
J Bactériol 168 : 702-707.
Google Scholar46.Dick RE, Quinn JP. 1995. Isolats dégradant le glyphosate provenant d’échantillons environnementaux : occurrence et voies de dégradation.
Appl Microbiol Biotechnologie 43 : 545-550.
Google Scholar47.Blake R, Pallett K. 2018. Le devenir environnemental et l’écotoxicité du glyphosate.
Google Scholar48.Imfeld G, Vuilleumier S. 2012. Mesurer les effets des pesticides sur les communautés bactériennes du sol : une revue critique.
Eur J Soil Biol 49 : 22-30.
Google Scholar49.Hart MM, Powell JR, Gulden RH, Dunfield KE, Peter Pauls K, Swanton CJ, Klironomos JN, Antunes PM, Koch AM, Trevors JT. 2009. Séparer l’effet de la culture de l’herbicide sur les communautés microbiennes du sol dans le maïs résistant au glyphosate.
Pédobiologie 52 : 253-262.
Google Scholar50.Laurence MH, Summerell BA, Burgess LW, Liew E. 2014. Concordance généalogique, reconnaissance des espèces phylogénétiques dans le complexe d’espèces
de Fusarium oxysporum .
Biol fongique 118 : 374-384.
Google Scholar51.Curran B, Jonas D, Grundmann H, Pitt T, Dowson CG. 2004. Développement d’un schéma de typage de séquences multilocus pour le pathogène opportuniste
Pseudomonas aeruginosa .
J Clin Microbiol 42 : 5644-5649.
Google Scholar52.Kepler RM, Rehner SA. 2013. Développement assisté par le génome de marqueurs de séquence nucléaire intergénique pour les champignons entomopathogènes du complexe d’espèces
Metarhizium anisopliae .
Mol Ecol Resour 13 : 210-217.
Google Scholar53.Kepler RM, Ugine TA, Maul JE, Cavigelli MA, Rehner SA. 2015. Composition de la communauté et génétique des populations de champignons pathogènes des insectes du genre
Metarhizium provenant des sols d’un système de recherche agricole à long terme.
Environ Microbiol 17 : 2791-2804.
Google Scholar54.Williams MM, Jr, Bradley CA, Duke SO, Maul JE, Reddy KN. 2015. L’incidence de la flétrissure de Goss dans le maïs sucré est indépendante des caractères transgéniques et du glyphosate.
HortScience 50 : 1791.
Google Scholar55.Brito IP, Tropaldi L, Carbonari CA, Velini ED. 2018. Effets hormétiques du glyphosate sur les plantes.
Pest Management Sci 74 : 1064-1070.
Google Scholar56.Duc SO. 2018. L’histoire et l’état actuel du glyphosate.
Pest Management Sci 74 : 1027-1034.
Google Scholar57.Duc SO, Reddy KN. 2018. La nutrition minérale des cultures résistantes au glyphosate est-elle altérée par le traitement au glyphosate ?
Outlook Pest Management 29 : 206-208.
Google Scholar58.Kremer R, Means N, Kim S. 2005. Le glyphosate affecte l’exsudation des racines de soja et les micro-organismes de la rhizosphère.
Int J Environ Anal Chem 85 : 1165-1174.
Google Scholar59.Cavigelli MA, Lengnick LL, Buyer JS, Fravel D, Handoo Z, McCarty G, Millner P, Sikora L, Wright S, Vinyard B, Rabenhorst M. 2005. Variation au niveau du paysage des ressources du sol et des propriétés microbiennes dans un maïs sans labour. champ.
Google Scholar60.Ross DS, Ketterings Q. 1995. Méthodes recommandées pour déterminer la capacité d’échange cationique du sol, p 62-70.
Dans Procédures d’analyse de sol recommandées pour le nord-est des États-Unis . Bulletin coopératif du Vermont no. 493.
Google Scholar61.EPA des États-Unis. 1986. Méthodes d’essai pour l’évaluation des déchets solides, 3e éd. SW-846. Service national d’information technique, Agence américaine de protection de l’environnement, Springfield, VA.
Google Scholar62.Komada H. 1975. Développement d’un milieu sélectif pour l’isolement quantitatif de
Fusarium oxysporum à partir du sol naturel.
Rév. Protection des Plantes Res 8 : 114-124.
Google Scholar63.Martin KJ, Rygiewicz PT. 2005. Amorces PCR spécifiques aux champignons développées pour l’analyse de la région ITS d’extraits d’ADN environnemental.
BMC Microbiol 5:28.
Google Scholar64.Caporaso JG, Lauber CL, Walters WA, Berg-Lyons D, Lozupone CA, Turnbaugh PJ, Fierer N, Knight R. 2011. Modèles globaux de diversité d’ARNr 16S à une profondeur de millions de séquences par échantillon.
Proc Natl Acad Sci USA 108 : 4516–4522.
Google Scholar65.White TJ, Bruns TD, Lee S, Taylor J. 1990. Amplification et séquençage direct des gènes d’ARN ribosomique fongique pour la phylogénétique, p 315-322.
Dans Innis MA, Gelfand DH, Sninsky JJ, White TJ (éd.),
Protocoles PCR, guide des méthodes et applications . Presse académique, San Diego, Californie.
Google Scholar66.Martin M. 2011. Cutadapt supprime les séquences d’adaptateur des lectures de séquençage à haut débit.
Google Scholar67.Callahan BJ, McMurdie PJ, Rosen MJ, Han AW, Johnson AJA, Holmes SP. 2016. DADA2 : inférence d’échantillons haute résolution à partir des données d’amplicons Illumina.
Méthodes Nat 13 : 581-583.
Google Scholar68.Wang Q, Garrity GM, Tiedje JM, Cole JR. 2007. Classificateur bayésien naïf pour l’attribution rapide de séquences d’ARNr dans la nouvelle taxonomie bactérienne.
Appl Environ Microbiol 73 : 5261–5267.
Google Scholar69.Nilsson RH, Larsson KH, Taylor AFS, Bengtsson-Palme J, Jeppesen TS, Schigel D, Kennedy P, Picard K, Glöckner FO, Tedersoo L, Saar I, Kõljalg U, Abarenkov K. 2019. La base de données UNITE pour l’identification moléculaire des champignons : gestion des taxons sombres et classifications taxonomiques parallèles.
Acides nucléiques Res 47 : D259–D264.
Google Scholar70.Quast C, Pruesse E, Yilmaz P, Gerken J, Schweer T, Yarza P, Peplies J, Glöckner FO. 2013. Le projet de base de données de gènes d’ARN ribosomal SILVA : traitement des données amélioré et outils Web.
Acides nucléiques Res 41 : D590–D596.
Google Scholar71.Dixon P. 2003. VEGAN, un ensemble de fonctions R pour l’écologie communautaire.
Google Scholar72.McMurdie PJ, Holmes S. 2013. Phyloseq : un package R pour une analyse interactive reproductible et des graphiques des données de recensement du microbiome.
Google Scholar73.Love MI, Huber W, Anders S. 2014. Estimation modérée du changement de pli et de la dispersion pour les données RNA-seq avec DESeq2.
Google Scholar74.McMurdie PJ, Holmes S. 2014. Ne gaspillez pas, ne voulez pas : pourquoi la raréfaction des données sur le microbiome est inadmissible.
Google Scholar75.Bokulich NA, Dillon MR, Zhang Y, Rideout JR, Bolyen E, Li H, Albert PS, Caporaso JG, Bokulich NA, Dillon MR, Zhang Y, Rideout JR, Bolyen E, Li H, Albert PS, Caporaso JG. 2018. q2-longitudinal : analyses longitudinales et par échantillons appariés des données du microbiome.
Impacts des systèmes de culture sur la teneur en glyphosate et en acide aminométhylphosphonique et sur la communauté microbienne des sols de grandes cultures au Québec (Canada)
L’utilisation d’herbicides à base de glyphosate (GBH) est omniprésente dans les grandes cultures du Québec, en dehors de la gestion biologique. Comme le glyphosate se dégrade généralement rapidement dans les sols agricoles, de l’acide aminométhylphosphonique (AMPA) est produit et persiste plus longtemps que le glyphosate. Des applications répétées de GBH année après année soulèvent des questions sur la pseudo-persistance du glyphosate et de l’AMPA dans les sols et sur leurs impacts possibles sur la communauté microbienne du sol. Cette recherche vise à comprendre l’influence des systèmes de culture et des propriétés édaphiques sur les teneurs en glyphosate et en AMPA ainsi que sur la diversité et la composition de la communauté microbienne du sol dans neuf champs de grandes cultures situés dans le sud du Québec (Canada) au cours des années 2019 et 2020. Teneurs moyennes du sol en glyphosate (0,16 ± 0,15 µg.g
−1 de sol sec) étaient inférieures à la teneur moyenne du sol en AMPA (0,37 ± 0,24 µg.g
−1 de sol sec). Les teneurs en glyphosate et en AMPA étaient significativement inférieures dans les sites cultivés en gestion biologique par rapport à la gestion conventionnelle. Pour les sites conventionnels, les doses cumulées de GBH ont eu un effet significatif sur la teneur en glyphosate du sol mesurée à la fin de la saison de croissance, mais pas sur la teneur en AMPA du sol. Les sites où les applications de GBH sont plus élevées semblent accumuler du glyphosate au fil du temps dans l’horizon du sol de 0 à 40 cm. La teneur en glyphosate et en AMPA du sol est inversement proportionnelle au pH du sol. La diversité alpha, la diversité bêta et le potentiel fonctionnel des communautés procaryotes et fongiques du sol n’ont pas été affectés par les doses cumulées de GBH, mais plutôt par les propriétés chimiques du sol, la texture du sol, la rotation des cultures et les apports de fumier.Mots clés:
La commercialisation de cultures résistantes au glyphosate au milieu des années 1990 a entraîné une augmentation importante de l’utilisation d’herbicides à base de glyphosate (GBH) [
1 ]. Le GBH peut être appliqué en pré- et post-levée sur les cultures résistantes au glyphosate [
2 ]. En 2021, le GBH était l’herbicide le plus utilisé représentant près de 50 % de tous les herbicides vendus au Québec, Canada [
3 ]. Le glyphosate a une affinité pour être adsorbé sur les particules du sol, bien qu’il puisse être transporté vers des profils de sol inférieurs en fonction des conditions météorologiques et des précipitations après l’application du GBH [
5 ]. La capacité d’adsorption du glyphosate sur les particules du sol est influencée par la capacité d’échange de cations (CEC), la texture du sol, la teneur en P et Al, la quantité de carbone organique du sol et le pH [
8 ]. Les molécules de glyphosate stérilisées qui ne s’adsorbent pas sur les tissus végétaux ou les particules du sol migreront dans les eaux interstitielles du sol, où le glyphosate est sujet à la dégradation [
4 ]. La dégradation du glyphosate est principalement biologique, bien que des voies abiotiques telles que la photolyse et la thermolyse puissent dégrader le glyphosate, produisant une variété de métabolites, dont l’acide aminométhylphosphonique (AMPA) [
10 ]. La biodégradation du glyphosate implique trois métabolites majeurs, l’AMPA, la sarcosine et l’acétyl-glyphosate [
11 ]. La voie qui produit l’AMPA est généralement favorisée par rapport aux deux autres voies dans les sols, avec environ 90 % des métabolites du glyphosate étant des molécules AMPA [
4 ]. La demi-vie du glyphosate se situe entre 1 et 197 jours, tandis que la demi-vie de l’AMPA se situe entre 23 et 958 jours dans les sols agricoles tempérés [
12 ]. Une fois dans le sol, l’AMPA est plus résistant à la biodégradation et a une affinité plus élevée pour les particules du sol que le glyphosate, ce qui augmente sa demi-vie dans le sol [
14 ]. Néanmoins, les deux molécules ont des propriétés physico-chimiques similaires avec une solubilité élevée dans l’eau, une faible lipophilie et un très faible potentiel de volatilisation [
16 ]. La principale différence entre les deux molécules est leur poids moléculaire (169,07 g·mol
−1 pour le glyphosate et 111,04 g·mol
−1 pour l’AMPA) [
17 ].L’agriculture de conservation gagne en popularité parmi les agriculteurs et les conseillers en cultures pour maintenir une productivité élevée des cultures tout en maintenant la communauté microbienne et les fonctions écosystémiques bénéfiques pour le sol, en réduisant les GES et en augmentant la séquestration du carbone et la résilience au changement climatique [
18 ]. Selon l’Organisation des Nations Unies pour l’alimentation et l’agriculture : « L’agriculture de conservation est un système agricole qui favorise une perturbation minimale du sol (c’est-à-dire sans labour), le maintien d’une couverture permanente du sol et la diversification des espèces végétales. Il améliore la biodiversité et les processus biologiques naturels au-dessus et au-dessous de la surface du sol, contribuant à une utilisation plus efficace de l’eau et des nutriments et à une production agricole améliorée et durable »[
19 ]. La production de grandes cultures au Québec, comme le maïs-grain, implique généralement des stratégies de désherbage mécanique ou chimique [
20 ]. Pour les producteurs adoptant les principes de l’agriculture de conservation tels que le semis direct et les cultures de couverture hivernales, le contrôle mécanique des mauvaises herbes est plus restrictif ou n’est pas recommandé, ce qui entraîne une dépendance à des doses d’application d’herbicides plus élevées pour contrôler les mauvaises herbes problématiques telles que les mauvaises herbes vivaces [
21 ]. L’utilisation de doses plus élevées de GBH pourrait poser problème en termes d’accumulation potentielle de glyphosate et d’AMPA dans les sols ou de dissipation dans les cours d’eau. Ceci est d’une grande pertinence puisque le glyphosate et l’AMPA ont été respectivement détectés dans 98,9 % et 93,3 % des échantillons d’eau des cours d’eau agricoles du Québec entre 2018 et 2020 [
22 ]. Le glyphosate et l’AMPA ont été détectés respectivement dans 42 % et 70 % des 45 sols agricoles québécois en 2014 [
23 ].Le séquençage à haut débit (HTS), également connu sous le nom de séquençage de nouvelle génération, a révolutionné l’étude des communautés microbiennes du sol. Comparé aux techniques telles que l’isolement basé sur la culture, l’électrophorèse sur gel à gradient dénaturant, l’électrophorèse sur gel à gradient de température, le polymorphisme de conformation simple brin et l’empreinte digitale par amplification de l’ADN, le HTS est à haut débit, moins cher et demande moins de main-d’œuvre [
24 ]. Avec HTS, l’identification taxonomique est effectuée sur la base de bases de données pour des parties spécifiques de gènes, par exemple, la région V4 de l’
27 ] pour les communautés procaryotes, fongiques et eucaryotes, respectivement. De plus, la réaction en chaîne par polymérase quantitative (qPCR) de régions spécifiques des gènes de l’ARNr pourrait être utilisée pour estimer la biomasse microbienne, tandis que la qPCR de gènes microbiens spécifiques pourrait également être utilisée pour quantifier des voies biologiques telles que la fixation du carbone [
30 ], qui peut demander beaucoup de travail lors de l’analyse de plusieurs gènes. L’avènement de bases de données telles que l’Encyclopédie des gènes et des génomes de Kyoto (KEGG) permet l’attribution directe du potentiel fonctionnel aux communautés microbiennes identifiées via HTS [
31 ]. Néanmoins, HTS présente des limites, basées sur la qualité de la base de données utilisée pour les identifications des potentiels taxonomiques et fonctionnels. Par exemple, l’identification taxonomique fongique est limitée en raison de la variabilité intraspécifique des séquences
ITS1 [
32 ].Des enquêtes antérieures sur la communauté microbienne du sol après l’application de GBH n’ont montré aucun effet sur la diversité alpha ou sur la composition globale [
36 ]. D’autre part, les techniques de culture utilisées dans l’agriculture de conservation, telles que le travail réduit du sol ou l’utilisation de cultures de couverture, sont connues pour augmenter considérablement la diversité alpha microbienne du sol et avoir un effet profond sur la composition de la communauté microbienne [
39 ]. Les effets interactifs des applications de GBH et des stratégies de systèmes de culture sur les communautés microbiennes du sol ont reçu peu d’attention jusqu’à présent [
36 ]. Jusqu’à présent, les études se sont concentrées sur les différences dans les applications GBH des années précédentes [
36 ] ou sur les différences dans le régime de travail du sol (labour direct ou travail au ciseau) [
33 ] ; cependant, ces études ont utilisé des parcelles expérimentales avec des systèmes de culture contrôlés, en se concentrant sur un paramètre à la fois. À notre connaissance, il n’existe aucune enquête dans les champs des producteurs commerciaux où ces facteurs (historique d’application d’herbicides, régime de travail du sol, utilisation des cultures de couverture) varient tous d’un producteur à l’autre.Pour mieux comprendre les effets interactifs des variables de gestion qui définissent les systèmes de culture, tels que la rotation des cultures, les applications de fumier, le travail du sol et les doses d’application de GBH, sur la teneur en glyphosate et en AMPA et sur la diversité des communautés microbiennes dans les sols, nous avons échantillonné les sols sur neuf champs de grandes cultures pendant deux années consécutives. Nous avons émis l’hypothèse que les teneurs en glyphosate et en AMPA seraient liées aux doses de GBH appliquées au cours des années précédentes et aux propriétés du sol telles que la CEC, le pH et la teneur en phosphore. De plus, nous avons émis l’hypothèse que la diversité (richesse et composition) et la diversité fonctionnelle de la communauté microbienne du sol seraient influencées par la combinaison des applications de GBH, des propriétés du sol et des pratiques de gestion des systèmes de culture.
2. Matériels et méthodes
2.1. Sélection et description des sites
Neuf champs avec une rotation, dont du maïs en 2019 et du soja en 2020, ont été sélectionnés dans le sud du Québec (Canada). Les régions de la Montérégie-Est et du Centre-du-Québec représentent respectivement 62 % et 15 % des superficies en grandes cultures de la province (
Figure 1 ). Deux champs (sites H et I) sont gérés par le même producteur. La sélection des champs visait à établir un gradient de systèmes de culture allant des systèmes biologiques sans application de GBH et labour conventionnel aux systèmes sans labour avec des doses variables de GBH appliquées.
Figure 1. Localisation des sites d’étude (
n = 9) dans la province de Québec, Canada. Un cercle rouge et une lettre jaune désignent chaque site.Après la sélection des sites, une enquête a été envoyée aux agriculteurs propriétaires des neuf sites. L’enquête comprenait des questions sur la fréquence et l’intensité du travail du sol, la stratégie de fertilisation organique et minérale, les applications de pesticides, les informations sur les semis et la récolte des grandes cultures et l’utilisation des cultures de couverture. À partir des informations recueillies lors des enquêtes, les neuf sites ont été divisés en différents groupes en fonction de la rotation des cultures utilisée, de la fréquence d’application du fumier, de l’intensité du travail du sol et des quantités cumulées de GBH appliquées au cours des 4 dernières années (
Figure 2 ). Les rotations de cultures ont été divisées en quatre catégories : deux rotations étaient constituées de maïs en alternance avec du soja, l’une en gestion biologique et l’autre en gestion conventionnelle. Une autre rotation incluait le maïs et le soja comme cultures principales, ainsi que le seigle céréalier comme culture de couverture entre la récolte et l’ensemencement. La quatrième rotation comprenait le maïs, le soja et le blé comme cultures principales ; un mélange de cultures de couverture a été semé après la récolte du blé. La fréquence du fumier était basée sur les applications effectuées entre 2017 et 2020. Deux applications ou plus ont été classées comme fréquentes, une comme peu fréquente et aucune application comme jamais. Pour les applications GBH, le nombre de grammes de glyphosate pulvérisé par hectare entre 2017 et 2020 a été compilé (
Tableau 1 ) et classé en deux groupes (>5 400 g·ha
-1 et <5 400 g·ha
-1 ).
Le tableau 2 présente les principales propriétés physico-chimiques des neuf sites.
Figure 2. Classification des neuf sites en fonction de la rotation des cultures, de la fréquence d’application du fumier, du travail du sol et de l’application cumulée de GBH entre 2017 et 2020. Acronymes : Org-2c : système biologique avec maïs et soja (MS) ; 2C : système MS classique ; 3C + CC : système conventionnel MS-Blé et cultures de couverture (céréales, radis, pois) après récolte du blé ; 2C + Cer Rye : système conventionnel avec MS avec culture de couverture (seigle céréalier) après chaque récolte.
Tableau 1. Informations sur les applications GBH sur les neuf sites.
Tableau 2. Propriétés physico-chimiques de chaque site pour le profil de sol 0–20 cm. Les valeurs moyennes sont présentées pour les échantillons de 2019 et 2020. Les valeurs
p proviennent d’une analyse de variance pour chaque variable ; les lettres proviennent d’une comparaison post-hoc par paire utilisant l’ajustement de Tukey.
2.2. Échantillonnage du sol
En septembre 2019 et 2020, chaque site a été échantillonné aux quatre mêmes emplacements géoréférencés. Ces emplacements représentaient l’extrémité d’un carré dont le côté mesurait 50 m. À chaque endroit, des carottes de sol ont été collectées à deux profondeurs (0 à 20 cm et 20 à 40 cm) à l’aide d’un carottier manuel de 8 cm de diamètre. Chaque échantillon consistait en un composite de quatre carottes de sol (total de ± 500 g) prélevées à l’extrémité d’un carré d’une longueur d’un mètre de côté. Après l’échantillonnage, tous les sols ont été homogénéisés, immédiatement mis sur la glace et transférés à -20 °C jusqu’à un traitement ultérieur.
2.3. Analyse du contenu du glyphosate et de l’AMPA
Les extractions de glyphosate et d’AMPA ont été réalisées selon l’approche décrite par Samson-Brais et al. [
40 ]. Les sols ont été lyophilisés et broyés à l’aide d’un pilier et d’un mortier. Cinq grammes de sol passés à travers un tamis de 2 mm ont été mélangés à 40 ml d’une solution d’extraction composée de 34,5 ml de NH
4 OH (28 à 30 %) (Fisher Scientific, Ottawa, ON, Canada) avec 13,6 g de KH
2 OREN
4 (Fisher Scientific, Ottawa, ON, Canada) dans un volume total de 1 L [
41 ]. La solution a été mélangée sur une roue tournante à 300 tr/min pendant 45 min suivie d’une centrifugation à 3 500 tr/min pendant 20 min. Ensuite, 40 µL du surnageant ont été transférés et évaporés à sec sous flux d’azote. Les échantillons ont ensuite été dérivés en utilisant 1 ml d’anhydride trifluoroacétique (TFAA) (Sigma-Aldrich, Saint-Louis, MO, USA) et 500 µL de trifluoroéthanol (TFE) (Sigma-Aldrich, Saint-Louis, MO, USA), suivis de chauffer pendant une heure à 100 °C. Après chauffage, les échantillons ont été refroidis à température ambiante, évaporés à sec sous flux d’azote puis remis en suspension avec 1 mL d’acétate d’éthyle avant injection (0,5 μL). Un chromatographe en phase gazeuse Varian CP 3800 couplé à un détecteur à capture d’électrons et équipé d’une colonne Rxi-5Sil MS (Restek, Bellefonte, PA, USA) (30 m × 0,25 mm × 0,25 μm) a été utilisé pour la quantification du glyphosate et de l’AMPA. L’injecteur et le détecteur ont été maintenus respectivement à 280 °C et 300 °C. De l’hydrogène a été utilisé comme gaz porteur avec un débit de colonne de 1,4 mL.min
-1 . Le programme de température du four a commencé à une température initiale de 70 °C, maintenue pendant 1 min, suivie d’une augmentation de 1 °C·min
−1 jusqu’à 84 °C, suivie d’une augmentation de 4 °C·min
−1 à 120 °C, puis suivi d’une augmentation de 80 °C·min
−1 jusqu’à 250 °C maintenue pendant 7 min, pour une durée totale de fonctionnement de 32,63 min.Les limites de détection (LOD) et de quantification (LOQ) calculées sont respectivement de 0,03 et 0,09 µg·g
−1 sol sec pour l’AMPA et de 0,02 et 0,05 µg·g
−1 sol sec pour le glyphosate [
23 ]. Les échantillons avec des valeurs inférieures à la LOD sont ajustés à 0,02 µg.g −1
de sol sec pour l’AMPA et 0,01 µg.g
−1 de sol sec pour le glyphosate, tandis que les échantillons avec des valeurs comprises entre la LOD et la LOQ sont ajustés à 0,06 µg.g
− 1 terre sèche pour AMPA et 0,03 µg·g
−1 sol sec pour le glyphosate. Pour quantifier le glyphosate et l’AMPA, une courbe d’étalonnage a été réalisée avec une matrice d’échantillon comprenant un blanc et cinq étalons différents pour chaque lot d’échantillons (0, 0,1, 0,2, 0,3, 0,4 et 0,6 μg·g
-1 et 0, 0,2 , 0,4, 0,6, 0,8 et 1,2 μg·g
−1 pour le glyphosate et l’AMPA, respectivement). Les courbes d’étalonnage ont montré une bonne linéarité (r
2 > 0,95 ;
p < 0,0001) dans la plage de concentrations attendue.
2.4. Analyse des propriétés physico-chimiques du sol
Les propriétés physiques (granulométrie, texture) et chimiques (C et N totaux, MO, pH, CEC, éléments Mehlich-3) (
Tableau 2 ) ont toutes été déterminées au laboratoire d’analyse agroenvironnementale de l’IRDA. La teneur en humidité du sol a été déterminée par séchage au four à 105 °C pendant 24 h. Les échantillons de sol composites ont été séchés à l’air, homogénéisés et tamisés à <2 mm. Le C total et le N total ont été déterminés sur des sous-échantillons finement broyés (100 mesh) par combustion sèche à l’aide d’un analyseur LECO-CN828. Le pourcentage de matière organique a été déterminé par perte au feu à 375 °C [
42 ]. Le pH du sol a été déterminé à l’aide d’une suspension sol:eau 1:1 [
43 ]. Une procédure similaire a été utilisée avec une suspension sol: solution SMP 1: 1 pour déterminer le pH du tampon du sol [
44 ]. Les concentrations de P, K, Ca, Mg, Al, B, Cu, Fe, Mn, Zn et Na ont été déterminées à l’aide d’un spectromètre d’émission optique à plasma à couplage inductif (ICP-OES, Perkin Elmer Optima 4300DV, Shelton, CT, USA ) après extraction Mehlich 3 [
45 ]. La CEC a été obtenue en calculant les cations basiques et acides en utilisant les résultats extractibles Mehlich-3 K, Ca, Mg et Na et le pH du tampon du sol [
46 ]. Le taux de saturation en phosphore (ISP) a été calculé en divisant le Mehlich-3 P extractible par le Mehlich-3 Al extractible [
2.5. Extraction d’ADN, séquençage et PCR quantitative d’ADN bactérien et fongique
Les extractions d’ADN ont été réalisées à l’aide d’un kit FastDNA Spin pour sol (MPBio, Irvine, CA, USA). Des échantillons de sol ont été ajoutés à des tubes contenant 1 mL du tampon de lyse et 1,4 g de la matrice de billes E fournie avec le kit. L’extraction de l’ADN a été réalisée conformément aux instructions du fabricant. Le culot d’ADN résultant a été mis en suspension dans 100 µL d’eau stérile de qualité moléculaire.La quantité et la qualité de l’extrait d’ADN ont été évaluées par spectrophotométrie à l’aide d’un biophotomètre (Eppendorf, Mississauga, ON, Canada) avec une cuvette G1,0 μ (Eppendorf, Mississauga, ON, Canada) avec des lectures à 260, 280, 230 et 320 nm. La région V4 du gène de
l’ARNr 16S des procaryotes (archées et bactéries) a été amplifiée à l’aide des amorces 515FB et 806RB [
48 ]. Pour les champignons, les eucaryotes (champignons)
ITS1 a été amplifié à l’aide des amorces BITS-ITS1 et B58S3 [
26 ]. Pour les communautés eucaryotes, le gène eucaryote de
l’ARNr 18S a été amplifié à l’aide des amorces E572F et B-E1009R [
27 ]. Tous les gènes ont été amplifiés dans le cadre d’une PCR à double index en deux étapes conçue pour les instruments Illumina par la Plateforme d’analyses génomiques (IBIS, Université Laval, Québec, QC, Canada).Le séquençage de l’ADN a été réalisé par IBIS sur une plateforme Illumina MiSeq, selon les méthodes de Jeanne et al. [
49 ]. Les procédures utilisées pour l’amplification et le séquençage de l’ADN fongique étaient similaires aux procédures utilisées pour l’amplification de l’ADN procaryote. Les séquences obtenues ont été démultiplexées en fonction du tag utilisé. Le contrôle de la qualité des séquences et la construction du tableau des caractéristiques ont été réalisés à l’aide de QIIME2 [
51 ]. Les bases de données de référence SILVA 138 [
52 ] (communautés procaryotes et eucaryotes) et UNITE version 8 [
53 ] (communauté fongique) ont été utilisées pour l’identification taxonomique des variantes de séquence d’amplicons [
54 ].Les quantités totales de bactéries et de champignons ont été quantifiées selon l’approche mentionnée dans [
55 ]. En bref, les paires d’amorces Eub-338/Eub518 et FF390/FR1 ont été utilisées respectivement pour les bactéries totales et les champignons totaux. À partir des valeurs CT moyennes des échantillons, les unités d’amplification ont été dérivées à l’aide de régressions linéaires conçues par le Laboratoire d’écologie microbienne de l’Institut de recherche et développement en agroenvironnement (IRDA, Québec, QC, Canada) comme décrit dans [
56 ].Pour évaluer le potentiel fonctionnel de la communauté procaryote dans les échantillons de sol, des approches d’inférence métabolique Picrust2 [
31 ] et les numéros de classification enzymatique de MetaCyc (EC) [
58 ]. Les enzymes et les voies métaboliques liées au glyphosate comprenaient le thiO (EC : 1.4.3.19) [
59 ], le phnP (EC : 3.1.4.55), le phnN (EC : 2.7.4.23), le phnM (EC : 3.6.1.63), le phnJ (EC :4.7.1.1) et phnIGHL (EC:2.7.8.37) [
60 ], et la voie du shikimate (M00022), qui sont tous liés à la dégradation du glyphosate dans le sol ainsi que la voie du shikimate connue pour être affectée par le glyphosate. Potentiels fonctionnels des voies générales telles que le métabolisme du soufre (M00176 ; M00596 ; M00595), le métabolisme des phosphonates (Ko00440), la nitrification (M00175 ; M00528 ; M00530 ; M00531 ; M00804), la dénitrification (M00529 ; M00973) et la fixation du carbone (M00165 à M00173). ; M00374 à M00377 ; M00579 ; M00620) ont été calculés. Des variants représentatifs de la séquence d’amplicons (ASV) ont été utilisés à partir de l’analyse QIIME2 sans filtration, suivis du pipeline par défaut avec des abondances de familles de gènes d’entrée non stratifiées par organismes contributeurs. Ces valeurs ont été normalisées en soustrayant la valeur moyenne de l’échantillon de la voie et en divisant par l’écart type de l’échantillon.
2.6. Analyse des données en aval et analyse statistique
Toutes les analyses statistiques ont été réalisées en R 4.2.2 [
61 ]. Les quantifications du glyphosate et de l’AMPA ont été visualisées dans le package ggplot2. Les contenus en glyphosate et en AMPA ont été transformés en log pour une analyse statistique. Les impacts de l’année et de la profondeur d’échantillonnage ainsi que des applications de GBH sur les contenus en glyphosate et en AMPA ont d’abord été évalués à l’aide d’une ANOVA, suivis d’une comparaison par paire post-hoc à l’aide de la fonction TukeyHSD du package de statistiques. Les différences entre les teneurs en glyphosate en 2019 et 2020 pour chaque lieu d’échantillonnage ont été utilisées pour étudier la dissipation de la molécule et la relier aux changements dans les applications de GBH entre 2019 et 2020. Une approche similaire a été utilisée pour comparer la dissipation de l’AMPA au changement dans Application de GBH entre 2019 et 2020. La teneur en glyphosate et en AMPA du sol en 2014 a été collectée auprès de Maccario et al. [
23 ]. Une ANOVA suivie de la fonction TukeyHSD a été réalisée sur des valeurs transformées en log pour évaluer l’effet des années sur la teneur en glyphosate et en AMPA. Des régressions multiples ont été utilisées pour évaluer l’effet des applications cumulatives de GBH entre 2017 et 2020, le pH du sol, la CEC, la teneur en argile et le rapport ISP sur la teneur en glyphosate et en AMPA. Une première série de régressions multiples a été réalisée avec les sites organiques et conventionnels, tandis qu’une deuxième série de régressions multiples a été réalisée avec uniquement les sites conventionnels.Pour les communautés procaryotes, fongiques et eucaryotes du sol, le nombre de lectures a été normalisé à 12 500, 13 000 et 3 900 lectures, respectivement, en utilisant la fonction raréfier-même-profondeur du package phyloseq R [
62 ]. La diversité alpha a été mesurée par les indices Shannon et Chao1 avec la fonctionestimate_richness du package phyloseq [
63 ]. L’impact des applications cumulatives de GBH et des systèmes de culture sur les indices de diversité microbienne et la quantification a été analysé à l’aide d’un modèle mixte avec un modèle de régression multiple, incluant l’année d’échantillonnage et la profondeur comme covariables fixes et le site comme effet aléatoire. La diversité bêta a été évaluée par analyse des coordonnées principales (PCoA) avec dissimilarité de Bray – Curtis pour une ordination non supervisée de la communauté microbienne du sol.
64 ]. Des analyses de redondance basées sur la distance (db-RDA) ont été utilisées pour évaluer l’effet marginal de toutes les propriétés physico-chimiques du sol sur la composition des communautés microbiennes. Les propriétés du sol ayant des effets marginaux significatifs ont été incluses dans une autre db-RDA avec des variables de gestion des sols.
3. Résultats
3.1. Contenu du sol en glyphosate et AMPA
Les teneurs en glyphosate du sol se situent entre en dessous de la LD et jusqu’à 0,72 µg·g
-1 de sol sec, tandis que les teneurs en AMPA dans le sol se situent entre en dessous de la LD et jusqu’à 1,22 µg·g
-1 de sol sec (
Tableau 3 ). Globalement, 16 % et 28 % des échantillons de sol présentent respectivement des teneurs en glyphosate inférieures à la LOD et comprises entre la LOD et la LOQ. Ensuite, 10 % et 28 % des échantillons de sol ont des teneurs en AMPA inférieures à la LOD et comprises entre la LOD et la LOQ, respectivement.
Tableau 3. Teneurs moyennes du sol en glyphosate et en AMPA en 2019 et 2020 par rapport aux catégories d’application cumulatives de GBH entre 2017 et 2020.
3.1.1. Impacts de l’année d’échantillonnage et de la profondeur d’échantillonnage
La teneur en glyphosate et en AMPA du sol est liée de manière significative à la profondeur d’échantillonnage, à l’année d’échantillonnage et aux applications cumulatives de GBH (
p < 0,01). L’interaction entre l’année d’échantillonnage et la profondeur est significative pour la teneur en glyphosate (
p < 0,01), alors qu’elle n’est pas significative pour l’AMPA (
p = 0,66) (
Tableau 3 ).Les teneurs en glyphosate du sol en 2020 à 20–40 cm de profondeur sont significativement inférieures (
p < 0,01) à celles de 0 à 20 cm de profondeur pour les deux années et à 20–40 cm de profondeur en 2019. Aucune autre différence significative n’est observée entre la profondeur d’échantillonnage et année. Les teneurs en glyphosate des sols des sites conventionnels sont significativement plus élevées que celles des sites biologiques (
p < 0,01). Plus précisément, les teneurs en glyphosate du sol en 2019 dans les exploitations ayant pulvérisé plus de 5 400 g·ha
−1 entre 2017 et 2020 sont plus élevées que celles des exploitations ayant pulvérisé moins de 5 400 g·ha
−1 (
p <0,01).Les teneurs en sol AMPA sont significativement plus élevées en 2019 par rapport à 2020 (
p < 0,01). Les teneurs en AMPA sont plus élevées entre 0 et 20 cm de profondeur que entre 20 et 40 cm de profondeur (
p < 0,01). Il y a beaucoup plus d’AMPA dans les sols des sites conventionnels que dans les sols biologiques (
p < 0,01). Cependant, la différence de teneur en AMPA entre les exploitations ayant pulvérisé plus de 5 400 g·ha
−1 et les exploitations ayant pulvérisé moins de 5 400 g·ha
−1 n’est pas significative (
p = 0,95).Les sites A, B, D, G, H&I sont gérés par des producteurs dont les champs ont déjà été échantillonnés en 2014 par Maccario et al. [
23 ]. Une comparaison entre la teneur en glyphosate et en AMPA de 2014, 2019 et 2020 a été effectuée (
Figure 3 ). Les sites organiques (A et B) ne présentent aucune augmentation ou diminution des teneurs en glyphosate et en AMPA. L’AMPA n’est pas significativement différent d’une année à l’autre pour tous les sites conventionnels, sauf entre 2014 et 2019 pour les sites H et I. La teneur en glyphosate est significativement plus élevée pour les sites G et H&I, mais pas significativement plus élevée pour le site D. Par conséquent, les sites avec des applications GBH plus élevées entre 2017 et 2020 semblent présenter une augmentation significative de la teneur en glyphosate entre 2014 et 2019-2020.
Figure 3. Teneurs du sol en glyphosate et AMPA dans les champs des producteurs ayant participé à la fois à cette expérience en 2019 et 2020 et à celle de Maccario et al. [
23 ] en 2014. Les sols ont été échantillonnés à une profondeur de 0 à 20 cm. Les lettres représentent une analyse post-hoc pour chaque molécule à l’aide de l’ajustement Tukey.Les changements dans la teneur en glyphosate du sol entre 2019 et 2020 dans la couche 0 à 20 cm sont corrélés aux changements dans les applications de GBH entre la moyenne de 2017 à 2019 et 2020 (
Figure S1 ). Cependant, cette corrélation n’est pas observée pour le glyphosate dans l’horizon 20-40 cm et l’AMPA dans l’horizon 0-20 cm. Les contenus en AMPA dans l’horizon de 20 à 40 cm présentent une corrélation négative avec l’application de GBH (
3.1.2. Impacts des pratiques de gestion et des propriétés du sol
Une première série de régressions multiples incluant des sites biologiques et conventionnels montre une corrélation significative entre les applications de GBH sur le glyphosate (
p < 0,01) et les teneurs en AMPA (
p < 0,01) dans l’horizon de sol de 0 à 20 cm. En revanche, toutes les propriétés physiques et chimiques du sol, à l’exception du pH du sol, n’ont pas de corrélation significative (
Figure S2 ). Cette corrélation des applications de GBH est due aux sites biologiques qui n’ont eu aucune application de GBH entre 2017 et 2020, ainsi qu’à de faibles teneurs en glyphosate et en AMPA. Après avoir exclu les sites organiques des régressions multiples, l’effet des applications de GBH est significatif pour le glyphosate (
p = 0,02) mais non significatif pour la teneur en AMPA (
Figure 4. Valeurs des coefficients ainsi que leur intervalle de confiance à 95 % pour toutes les variables indépendantes dans les régressions multiples pour la teneur en AMPA (
à droite ) et en glyphosate (
à gauche ) pour un horizon de sol de 0 à 20 cm de sites conventionnels.En excluant les sites organiques des régressions multiples, les valeurs R carré ajustées des régressions sont de 50 % pour le glyphosate et de 19 % pour l’AMPA. Le pH du sol montre un effet négatif significatif sur la teneur en glyphosate (
p < 0,01) et en AMPA (
p < 0,01). La teneur en argile du sol montre un effet positif sur la teneur en glyphosate (
p = 0,03) mais pas sur la teneur en AMPA (
p = 0,59). La CEC du sol n’a pas d’effet significatif sur le glyphosate (
p = 0,22), mais un effet positif significatif sur la teneur en AMPA (
p = 0,02). L’ISP du sol a un effet négatif significatif sur la teneur en glyphosate (
p = 0,01) mais pas sur la teneur en AMPA (
p = 0,91).
3.2. Communauté microbienne du sol
Le séquençage des amplicons sur MiSeq des trois groupes microbiens a donné, après filtration, une moyenne de 27 788 (± 4 818) ; 28 740 (±7 809) ; et 4 819 (± 1 464) séquences par échantillon pour les procaryotes, les champignons et les eucaryotes, respectivement. En combinant tous les échantillons, la communauté procaryote contenait 17 400 ASV uniques, la communauté fongique contenait 17 500 ASV uniques et la communauté eucaryote contenait 6 000 ASV uniques.
3.2.1. Alpha-Diversité et Abondance
L’analyse des mesures de diversité alpha montre une forte corrélation entre Shannon, Chao1, les ASV observés et les mesures d’uniformité pour chaque communauté microbienne. Par conséquent, l’indice de diversité de Shannon a été utilisé pour évaluer la diversité alpha des communautés microbiennes, tandis que les résultats de quantification ont évalué l’abondance totale des procaryotes et des champignons (
Figure 5 ). L’indice de Shannon pour les communautés fongiques et eucaryotes n’est pas significativement affecté par la profondeur d’échantillonnage (
p > 0,05) ou l’année (
p > 0,10). En revanche, l’indice de Shannon procaryote est significativement impacté par l’année d’échantillonnage (
p < 0,001) avec une diversité plus élevée en 2020 par rapport à 2019, mais aucun effet significatif de la profondeur d’échantillonnage (
p = 0,19). La quantification des communautés procaryotes et fongiques est significativement influencée par l’année d’échantillonnage (
p < 0,001) et la profondeur d’échantillonnage (
p < 0,001). Pour les deux communautés microbiennes, les quantités totales sont plus élevées dans la couche de 0 à 20 cm. Pour les communautés procaryotes, la quantification est plus élevée en 2019 sur les sites A, B, C, D, G et H, tandis qu’elle est plus élevée pour les communautés fongiques en 2020 sur les sites C, D, F, G, H, I.
Figure 5. Valeur normalisée de l’indice de diversité de Shannon des communautés procaryotes (Shannon_Prok), fongiques (Shannon_Fung) et eucaryotes (Shannon_Euk) ainsi que quantification du total des bactéries (Total_Prok) et du total des champignons (Total_Fung) pour chaque site pour les années 2019 (maïs) et 2020 (soja) et les deux horizons (0-20 cm et 20-40 cm).Les applications cumulatives de GBH et la gestion des cultures ont un faible effet sur la diversité et la richesse des communautés microbiennes. La diversité de la communauté procaryote et fongique de Shannon n’est pas significativement affectée par les applications cumulatives de GBH (
p > 0,27) et les rotations des cultures (
p > 0,05). Les applications GBH ont un impact significatif sur la diversité eucaryote (
p = 0,02), avec une diversité eucaryote plus élevée dans les domaines recevant des applications GBH. Les applications cumulatives de GBH et les rotations des cultures n’ont pas d’impact sur la quantification des procaryotes et des champignons.
3.2.2. Bêta-Diversité
La composition des communautés microbiennes, notamment procaryotes, est fortement influencée par la texture du sol (
Figure 6 ). L’analyse PCoA montre que les communautés microbiennes trouvées dans les sols à texture grossière ont tendance à différer de celles des sols à texture fine, avec des différences marquées pour les communautés procaryotes et, dans une moindre mesure, pour les communautés fongiques et eucaryotes. Il est intéressant de noter que les communautés fongiques du sol à texture fine couvrent une grande partie de la parcelle d’ordination, ce qui indique que les communautés fongiques dans un sol à texture fine peuvent être très diverses par rapport aux communautés procaryotes, qui sont plus similaires dans un sol à texture fine (
Figure 6. Analyse des coordonnées principales des communautés procaryotes (
à gauche ), fongiques (
au milieu ) et eucaryotes (
à droite ). Les couleurs des points représentent les groupes basés sur les applications cumulatives GBH entre 2017 et 2020. Les formes de points et les couleurs des ellipses représentent la texture du sol. Le seuil de signification des ellipses était de 0,05.À l’aide de db-RDA, un ensemble de variables de propriétés du sol (MO, Ca, Na, teneur en argile et teneur en sable) ont été identifiées comme ayant un effet significatif sur la composition des trois communautés microbiennes (
tableau supplémentaire S1 ). L’effet marginal de ces propriétés du sol sélectionnées est comparé à l’effet marginal des pratiques de gestion (
Tableau 4 ). Dans l’ensemble, la concentration en sodium a l’effet marginal le plus élevé pour toutes les communautés microbiennes, suivie par la teneur en argile et en sable, puis par la rotation des cultures, les apports de fumier et la teneur en calcium. Bien que la teneur en matière organique ait un effet marginal significatif lorsque l’on considère uniquement les variables environnementales (
Tableau supplémentaire S1 ), l’effet n’est pas significatif dans un modèle incluant des variables de gestion (
Tableau 4 ). Les doses d’application de GBH n’ont pas d’effet marginal significatif sur les communautés microbiennes du sol.
Tableau 4. Effet marginal de l’analyse de redondance basée sur la distance de chaque variable environnementale et de gestion sur les communautés microbiennes du sol.
3.2.3. Potentiel fonctionnel
Des outils tels que Picrust2 permettent de déduire différents potentiels fonctionnels, tels que les gènes impliqués dans la dégradation du glyphosate, ou des voies générales qui sont des processus importants dans le sol à partir des communautés microbiennes observées dans notre expérience (
Figure 7 ). Dans l’ensemble, l’année d’échantillonnage a un effet très significatif (
p < 0,001) sur tous les gènes et voies d’intérêt, avec des valeurs plus élevées en 2020 par rapport à 2019. L’horizon du sol n’a pas d’effet significatif pour tous les gènes, à l’exception des gènes liés. à l’opéron phn (phnIGHL, phnJ, phnM, phnN et phnP), pour lequel le potentiel était plus élevé dans l’horizon 0-20 cm que dans l’horizon 20-40 cm.
Figure 7. Potentiel fonctionnel normalisé des communautés procaryotes pour les gènes liés au métabolisme général (
en haut ) et les gènes liés au métabolisme du glyphosate (
en bas ) pour les deux années (2019 et 2020) et les deux horizons du sol (0-20 cm et 20-40 cm) .La texture du sol, exprimée en teneur en sable et en argile, a le plus d’effets à la fois sur les fonctions générales et sur les fonctions liées au glyphosate. Les rotations de cultures et les apports de fumier montrent un effet significatif sur l’opéron phn, à l’exception du phnP. Les apports fréquents de fumier ont un potentiel de phnM significativement plus élevé (
p = 0,044) par rapport aux apports peu fréquents de fumier. Les applications cumulatives GBH n’ont pas d’effet significatif sur les fonctions d’intérêt.
4. Discussion
4.1. Pseudo-persistance du glyphosate et de l’AMPA dans les sols
La pseudo-persistance des polluants est définie par l’ajout constant de nouvelles molécules qui reconstituent les molécules éliminées [
65 ]. Comme les GBH sont appliqués presque chaque année sur des sols qui ne sont pas sous gestion biologique, le glyphosate et l’AMPA pourraient être considérés comme des polluants pseudo-persistants dans ces sols [
66 ]. La pseudo-persistance du glyphosate et de l’AMPA dans les sols échantillonnés analysés est presque omniprésente, avec 84 % des échantillons contenant des niveaux détectables de glyphosate et 90 % pour l’AMPA. Les sites sans application de GBH depuis plus d’une décennie présentent des niveaux détectables de glyphosate et d’AMPA dans leurs sols. Les traces de glyphosate et d’AMPA dans les sites biologiques peuvent s’expliquer par l’épandage fréquent de fumier sur les champs biologiques. Comme le fumier ne doit pas nécessairement provenir d’animaux cultivés de manière biologique, il peut contenir des résidus de glyphosate et d’AMPA provenant de l’alimentation animale conventionnelle [
67 ] a épandu du fumier de volaille sur un champ qui n’a jamais reçu de GBH. Avec un taux d’épandage de fumier extrêmement élevé (36 MT·ha
−1 ), Fuchs et al. ont pu détecter 1,7 µg·g
−1 de sol sec de glyphosate dans un sol organique. Cette teneur en glyphosate est beaucoup plus élevée que toutes les mesures rapportées ici dans les champs biologiques et conventionnels, démontrant la capacité du fumier à importer du glyphosate et de l’AMPA dans les champs gérés biologiquement.Des expériences antérieures axées sur les sols de grandes cultures rapportent des niveaux de détection de glyphosate et d’AMPA légèrement inférieurs ou similaires dans les sols du Québec (42 % et 70 %), de l’Argentine (100 % pour les deux) et du Brésil (94 % et 100 %). respectivement [
68 ]. Le taux de détection plus faible enregistré au Québec a été observé dans les échantillons de sol prélevés en 2014 dans les champs de soja [
23 ]. Toutes ces études ont été réalisées sur des sols avec des applications fréquentes de GBH dans la production de maïs et de soja, expliquant l’omniprésence du glyphosate et de l’AMPA. Bien que les taux de détection soient similaires dans toutes les régions du monde, nos résultats suggèrent que les sols de grandes cultures non biologiques du Québec qui ont été échantillonnés dans cette étude contiennent légèrement plus de glyphosate (0,17 µg·g
−1 sol sec ±0,15) et d’AMPA (0,37 µg·g· g
−1 sol sec ±0,24) par rapport aux sols brésiliens (0,08 µg·g
−1 sol sec ±0,09 pour le glyphosate et 0,17 µg·g
−1 sol sec ±0,16 pour l’AMPA) et européens (0,11 µg·g
69 ] bien que plus petit que le sol argentin (2,30 µg·g
−1 sol sec ±0,48 et 4,20 µg·g
−1 sol sec ±2,26) [
66 ]. Les teneurs en glyphosate et en AMPA mesurées dans l’horizon 0-20 cm en 2019-2020 sont généralement supérieures à celles de l’étude de Maccario menée en 2014 (0,06 µg·g
−1 sol sec ±0,10 et 0,29 µg·g
−1 sol sec ±0,21) [
23 ]. Notre analyse montre que les contenus en AMPA ne sont pas significativement différents entre 2014 et 2020 pour cinq producteurs. En revanche, il existe une différence significative de teneur en glyphosate entre 2014 et 2020 pour les deux producteurs ayant des demandes de GBH supérieures à 5 400 g·ha
−1 entre 2017 et 2020. Ce constat est en accord avec Primost et al. [
66 ] qui ont rapporté que la teneur en glyphosate était mieux corrélée aux applications cumulées de GBH sur plusieurs années par rapport à la dernière dose de pulvérisation. L’accumulation de glyphosate dans le sol semble se produire à un rythme inférieur à celui prévu par Primost et al. [
66 ]. Il a été suggéré qu’une augmentation de la teneur en glyphosate de 1 µg·g
−1 de sol sec se produirait toutes les 5 applications de GBH [
66 ]. Cependant, au cours de plus de 6 années d’applications annuelles de GBH, nous avons observé une augmentation moyenne de 0,20 µg·g
−1 de sol sec pour trois champs (sites G, H, I) et aucune augmentation significative pour un champ (site D). ).
4.2. Impacts des propriétés du sol et des systèmes de culture sur la teneur en glyphosate et en AMPA
Nos résultats démontrent que la gestion biologique entraîne une différence significative dans la teneur en glyphosate et en AMPA dans les sols, ce qui est intuitif puisque l’application de GBH est interdite dans ce type de système de culture (
Figure S2 ). Lorsque l’on exclut les sites biologiques de notre analyse, les applications cumulatives de GBH trois ans avant l’échantillonnage influencent de manière significative la teneur en glyphosate du sol, mais pas la teneur en AMPA (
Figure 4 ). Cette différence entre le comportement du glyphosate et celui de l’AMPA dans les sols pourrait s’expliquer par la pseudo-persistance du glyphosate liée à des apports fréquents de glyphosate supérieurs aux taux de dissipation [
66 ] et par l’utilisation de résidus de culture comme couverture de sol pour le semis direct. chantiers [
70 ]. Même avec un taux de dissipation rapide du glyphosate dans les sols [
71 ], des apports relativement élevés de glyphosate (> 1 350 g·ha
-1 ·an
-1 ) dans les sols agricoles pourraient entraîner une accumulation partielle de glyphosate qui ne se dégrade pas dans l’AMPA et lessivage supplémentaire dans les horizons plus profonds du sol, en particulier après de fortes pluies [
72 ]. Pour les sols recouverts de biomasse végétale (c’est-à-dire des plantes vivantes ou des résidus de cultures) au moment de l’application de GBH, une partie du GBH appliqué est interceptée par la biomasse végétale plutôt que sur le sol, ce qui modifie la dissipation du glyphosate [
75 ]. Le glyphosate intercepté par la biomasse végétale ralentira généralement la dissipation du glyphosate par rapport à l’application directement sur le sol, car une plus grande partie du glyphosate est incorporée ou adsorbée sur la biomasse végétale par rapport aux particules du sol [
73 ]. La biomasse végétale déjà partiellement dégradée adsorbera le glyphosate plus fortement que la biomasse végétale fraîche [
73 ] et une plus grande partie du glyphosate restera sous forme de résidus non extractibles trouvés dans la biomasse végétale [
75 ]. Le recours au travail du sol pour incorporer les résidus de culture accélérera la dégradation de la biomasse végétale en la mélangeant au sol [
70 ]. Par conséquent, les sites avec des pratiques sans labour et une utilisation fréquente de cultures de couverture (sites G, H et I) pourraient entraîner une dissipation plus lente du glyphosate par rapport aux sites avec un travail du sol et une couverture du sol plus faible par les résidus de culture (site D). De plus, des applications répétées de GBH peuvent entraîner une diminution de l’activité microbienne du sol et de la respiration du sol [
77 ], inhibant potentiellement la biodégradation du glyphosate en AMPA [
40]. Par conséquent, des applications plus élevées de GBH ne pourraient pas conduire directement à des teneurs plus élevées en AMPA dans le sol, mais plutôt à une accumulation de glyphosate. De telles interprétations pourraient expliquer la teneur plus élevée en glyphosate dans les profils de sol plus profonds pour les sites avec > 5 400 g·ha
−1 de GBH appliqués entre 2017 et 2020 (
Tableau 3 ) et l’augmentation avec le temps du glyphosate dans les mêmes sites échantillonnés en 2014 et en 2020. (
Figure 3 ).Le pH du sol, l’ISP et la teneur en argile ont un effet significatif sur la teneur en glyphosate et en AMPA du sol, similaire aux expériences précédentes [
8 ]. Nos observations d’une relation négative générale entre le glyphosate et le pH du sol et d’une relation positive entre le glyphosate et la teneur en argile sont en accord avec la littérature [
81 ]. Une étude réalisée en Argentine a montré que le glyphosate présente une affinité plus élevée pour les surfaces du sol à un certain pH, qui varie selon les types de sol mais se situe généralement autour du pH 6 [
82 ], ce qui correspond aux valeurs de pH du sol plus faibles enregistrées dans nos expériences. Dans la même étude, il a été démontré que la présence de phosphate réduit l’adsorption du glyphosate sur les particules du sol [
82 ], ce qui concorde avec notre découverte d’une relation négative entre la teneur en glyphosate et la valeur ISP du sol (
Figure 4 ). La teneur en argile et l’ISP ne présentent pas de corrélation significative avec les teneurs en AMPA, tandis que la CEC du sol a une relation positive avec les teneurs en AMPA. Maccario et coll. [
23 ] a également constaté que la texture du sol n’avait pas d’impact sur la teneur en AMPA. Cela pourrait s’expliquer par la plus grande affinité du sol pour l’AMPA par rapport au glyphosate [
4.3. Impacts des systèmes de culture et des propriétés du sol sur la communauté microbienne du sol
La diversité alpha et la biomasse totale des communautés procaryotes et fongiques ne sont pas affectées par les rotations des cultures dans notre étude. Seule la diversité alpha eucaryote est significativement impactée par les applications cumulatives du GBH. Les applications cumulatives élevées de GBH dans cette étude sont liées à d’autres choix de gestion tels que la culture sans labour. Par conséquent, la relation positive entre l’application du GBH et les communautés eucaryotes pourrait s’expliquer par l’adoption du semis direct ainsi que par une application élevée du GBH. En effet, Kepler et al. [
33 ] ont montré que l’impact des applications de GBH sur les communautés microbiennes du sol était négligeable par rapport aux pratiques de travail du sol. Bien qu’une méta-analyse n’ait montré aucun impact significatif sur la diversité fongique des pratiques sans labour [
84 ], d’autres études ont montré l’effet négatif du travail du sol sur la communauté eucaryote du sol [
88 ], ce qui conforte notre hypothèse.La composition de la communauté microbienne du sol et son potentiel fonctionnel sont influencés à la fois par les propriétés du sol et les systèmes de culture dans notre étude. La teneur en sodium a l’effet le plus important sur la composition des communautés microbiennes, ce qui est difficile à expliquer. Le sodium du sol peut avoir des effets négatifs importants à des teneurs élevées (1 000 ppm) [
90 ] ; cependant, dans cette étude, tous les échantillons de sol présentent des teneurs en Na inférieures à 40 ppm avec des valeurs aussi basses que 2,7 ppm. Aucun effet important du Na sur les communautés microbiennes du sol n’a été signalé aux concentrations observées dans notre étude. La texture du sol a un effet important sur la composition des communautés microbiennes et leur potentiel fonctionnel. De nombreuses études ont confirmé l’effet déterminant de la texture du sol sur la composition et le potentiel fonctionnel des communautés microbiennes [
93 ]. En termes de systèmes de culture, la rotation des cultures et la fréquence d’application du fumier sont souvent citées comme ayant un effet significatif sur les communautés microbiennes du sol [
95 ] ont montré que l’application d’engrais, en particulier de fumier, avait un rôle plus important que la rotation des cultures et le stade de croissance des cultures sur les communautés microbiennes du sol. Une autre étude a montré que toutes les communautés microbiennes ne sont pas affectées de la même manière par la rotation des cultures, les communautés fongiques étant plus sensibles au nombre d’espèces végétales dans une rotation des cultures par rapport aux communautés procaryotes [
97 ]. Dans la présente étude, aucun effet significatif de l’application de GBH sur la structure de la communauté microbienne du sol et son potentiel fonctionnel n’a été observé, ce qui est cohérent avec d’autres rapports de la littérature utilisant des approches ADN pour évaluer les communautés microbiennes [
99 ]. Cela suggère que d’autres pratiques de gestion, telles que la rotation des cultures, les apports de fumier et le travail du sol, ont un impact plus important sur la composition de la communauté microbienne du sol et sur son potentiel fonctionnel que les applications GBH.
5. Conclusions
Le glyphosate et l’AMPA ont été détectés dans les sols de tous les sites étudiés, y compris les sites biologiques. Les doses cumulées de GBH ne sont pas corrélées au contenu du sol AMPA dans les sites conventionnels. Cependant, la teneur en glyphosate du sol était positivement corrélée aux doses cumulées de GBH, et il semble y avoir eu une augmentation de la teneur en glyphosate du sol entre 2014 et 2020 pour les sites ayant les applications de GBH les plus importantes. Un faible pH du sol et une teneur élevée en argile correspondaient à une pseudo-persistance plus élevée du glyphosate et de l’AMPA dans le sol. Les quantités cumulatives d’applications de GBH ne semblent pas avoir d’impact sur les communautés microbiennes du sol, tandis que la rotation des cultures et les apports de fumier ont un impact significatif. Par conséquent, les applications de GBH semblent avoir un impact minimal sur les communautés microbiennes du sol et sur le contenu du sol AMPA. D’un autre côté, les applications de GBH semblent augmenter la teneur en glyphosate du sol et doivent être surveillées pour garantir que l’accumulation de glyphosate n’entraîne pas d’effets néfastes sur les communautés microbiennes du sol.
Matériel supplémentaire
Les informations complémentaires suivantes peuvent être téléchargées sur :
https://www.mdpi.com/article/10.3390/agronomy14040686/s1 , Figure S1 : Corrélation entre la différence de teneur en glyphosate (en haut) et en AMPA (en bas) entre 2019 et 2020 et la différence dans l’application de GBH en 2020 et l’application annuelle moyenne de GBH entre 2017 et 2019. Figure S2 : valeurs des coefficients ainsi que leur intervalle de confiance à 95 % pour toutes les variables indépendantes dans des régressions multiples pour la teneur en AMPA (à droite) et en glyphosate (à gauche) pour 0– Profil de sol de 20 cm de tous les sites. Figure S3 : Chromatographes d’une courbe d’étalonnage en six points pour l’AMPA (A) et d’une courbe d’étalonnage en cinq points pour le glyphosate (B) sur une matrice de sol. Tableau supplémentaire S1 : Effet marginal des propriétés du sol sur la composition des communautés microbiennes pour l’horizon 0–20 du sol.
Contributions d’auteur
Conceptualisation, RH, JD-P., TJ, MM et ML ; méthodologie, WO, RH, JD-P., TJ, MM et ML ; validation, WO, JD-P. et MM ; analyse formelle, WO ; enquête, adjudant, CP et JD-P.; conservation des données, WO et CP ; rédaction – préparation de l’ébauche originale, WO ; rédaction – révision et édition, WO, RH, JD-P., TJ, MM et ML ; visualisation, WO ; supervision, RH et ML ; administration de projet, RH ; acquisition de financement, RH et ML Tous les auteurs ont lu et accepté la version publiée du manuscrit.
Financement
Le financement de cette recherche provenait du programme Innov’Action Agroalimentaire, dans le cadre du Partenariat canadien pour l’agriculture, une entente entre les gouvernements du Canada et du Québec.
Déclaration de disponibilité des données
Des ensembles de données accessibles au public ont été analysés dans cette étude. Ces données et scripts de figures peuvent être trouvés ici :
Les auteurs souhaitent remercier tous les agriculteurs qui nous ont permis d’échantillonner leurs champs et ont donné de leur temps pour répondre à nos questions.
Les conflits d’intérêts
Les auteurs ne déclarent aucun conflit d’intérêts.
Les références
Novotny, E. Glyphosate, Roundup et les échecs de l’évaluation réglementaire. Toxiques 2022 , 10 , 321. [ Google Scholar ] [ CrossRef ] [ PubMed ]
Szekacs, A. ; Darvas, B. Quarante ans avec le glyphosate. Herbique.-Prop. Synthé. Contrôler les mauvaises herbes 2012 , 14 , 247-284. [ Google Scholar ]
Ministère de l’Environnement, de la Lutte Contre les Changements Climatiques. Bilan des Ventes de Pesticides au Québec—Année 2021 ; Bibliothèque et Archives Nationales du Québec : Montréal, QC, Canada, 2023. [ Google Scholar ]
Kanissery, R. ; Gairhe, B. ; Kadyampakeni, D. ; Batuman, O. ; Alferez, F. Glyphosate : Sa persistance dans l’environnement et son impact sur la santé et la nutrition des cultures. Plantes 2019 , 8 , 499. [ Google Scholar ] [ CrossRef ] [ PubMed ]
Carretta, L. ; Cardinali, A. ; Onofri, A. ; Masin, R. ; Zanin, G. Dynamique du glyphosate et de l’acide aminométhylphosphonique dans le sol sous labour conventionnel et de conservation. Int. J. Environ. Rés. 2021 , 15 , 1037-1055. [ Google Scholar ] [ CrossRef ]
Motharasan, M. ; Yunus ; Shukor, Y. ; Yasid, N. ; Wan Johari, WL ; Johari, W. ; Aqlima, S. Devenir environnemental et dégradation du glyphosate dans le sol. Pertanika J. Sch. Rés. Rév.2018 , 4 , 102-116 . [ Google Scholar ]
Dollinger, J. ; Dagès, C. ; Voltz, M. Adsorption du glyphosate dans les sols et les sédiments prédite par les fonctions de pédotransfert. Environ. Chimique. Lett. 2015 , 13 , 293-307. [ Google Scholar ] [ CrossRef ]
De Gerónimo, E. ; Aparicio, VC; Costa, JL Adsorption du glyphosate dans les sols d’Argentine. Estimation du coefficient d’affinité par fonction de pédotransfert. Géoderme 2018 , 322 , 140-148. [ Google Scholar ] [ CrossRef ]
Ascolani Yael, J. ; Fuhr, JD; Bocan, Géorgie ; Daza Millone, A. ; Tognalli, N. ; dos Santos Afonso, M. ; Martiarena, ML Dégradation abiotique du glyphosate en acide aminométhylphosphonique en présence de métaux. J. Agric. Chimie alimentaire. 2014 , 62 , 9651-9656. [ Google Scholar ] [ CrossRef ]
Papagiannaki, D. ; Médana, C. ; Binetti, R. ; Calza, P. ; Roslev, P. Effet de l’irradiation UV-A, UV-B et UV-C du glyphosate sur la photolyse et l’atténuation de la toxicité aquatique. Sci. Rep. 2020 , 10 , 20247. [ Google Scholar ] [ CrossRef ]
Singh, S. ; Kumar, V. ; Gill, JPK; Datta, S. ; Singh, S. ; Dacca, V. ; Kapoor, D. ; Wani, Alberta; Dhanjal, DS; Kumar, M. ; et coll. Herbicide Glyphosate : toxicité et dégradation microbienne. Int. J. Environ. Rés. Santé publique 2020 , 17 , 7519. [ Google Scholar ] [ CrossRef ]
Carretta, L. ; Masin, R. ; Zanin, G. Revue des études analysant la présence de glyphosate et d’acide aminométhylphosphonique (AMPA) dans les eaux souterraines. Environ. Rév.2021 , 30 , 88-109 . [ Google Scholar ] [ CrossRef ]
Tang, FHM ; Jeffries, TC ; Vervoort, RW; Conoley, C. ; Coleman, Nevada ; Maggi, F. Expériences de microcosme et modélisation cinétique de la biodégradation du glyphosate dans les sols et les sédiments. Sci. Environ. Total. 2019 , 658 , 105-115. [ Google Scholar ] [ CrossRef ]
Wimmer, B. ; Langarica-Fuentes, A. ; Schwarz, E. ; Kleindienst, S. ; Huhn, C. ; Pagel, H. La modélisation mécanistique indique une dissipation rapide du glyphosate et une persistance induite par la sorption de son métabolite AMPA dans le sol. J. Environ. Qual. 2023 , 52 , 393-405. [ Google Scholar ] [ CrossRef ]
Blake, R. ; Pallett, K. Le devenir environnemental et l’écotoxicité du glyphosate. Gestion antiparasitaire Outlook. 2018 , 29 , 266-269. [ Google Scholar ] [ CrossRef ]
von Mérey, G. ; Manson, PS ; Mehrsheikh, A. ; Sutton, P. ; Levine, SL Évaluation des risques chroniques pour le glyphosate et l’acide aminométhylphosphonique pour le biote du sol. Environ. Toxicol. Chimique. 2016 , 35 , 2742-2752. [ Google Scholar ] [ CrossRef ]
Kim, YJ ; Nitin, N. ; Kim, KB Différences toxicocinétiques négligeables du glyphosate par différents véhicules chez le rat. Toxiques 2023 , 11 , 67. [ Google Scholar ] [ CrossRef ]
Kassam, A. ; Friedrich, T. ; Derpsch, R. Propagation mondiale de l’agriculture de conservation. Int. J. Environ. Goujon. 2019 , 76 , 29-51. [ Google Scholar ] [ CrossRef ]
Bilodeau, L.; Marcoux, A. ; Miville, D. ; Fréchette, I. ; Samson, V. Désherbage du Maïs de Grandes Cultures. 2020. Disponible en ligne : https://www.agrireseau.net/documents/Document_100450.pdf (consulté le 20 février 2024).
Kumar, V. ; Mahajan, G. ; Dahiya, S. ; Chauhan, BS Défis et opportunités pour la gestion des mauvaises herbes dans les systèmes agricoles sans labour. Dans Systèmes de culture sans labour pour une agriculture durable : défis et opportunités ; Dang, YP, Dalal, RC, Menzies, NW, éd. ; Springer International Publishing : Cham, Suisse, 2020 ; pp. 107-125. [ Google Scholar ]
Giroux, I. Présence de Pesticides dans l’Eau au Québec : Portrait et Tendances dans les Zones de Maïs et de Soya—2018 à 2020 ; Ministère de l’Environnement et de la Lutte Contre les Changements Climatiques, Direction de la Qualité des Milieux Aquatiques : Québec, QC, Canada, 2022. [ Google Scholar ]
Maccario, S. ; Lucotte, M. ; Moingt, M. ; Samson-Brais, É.; Smedbol, É. ; Labrecque, M. Impact des caractéristiques des sols et des pratiques de gestion des mauvaises herbes sur la persistance du glyphosate et de l’AMPA dans les sols de grandes cultures des basses terres du Saint-Laurent (Québec, Canada). Agronomie 2022 , 12 , 992. [ Google Scholar ] [ CrossRef ]
Nkongolo, KK; Narendrula-Kotha, R. Progrès dans la surveillance de la dynamique et de la fonction de la communauté microbienne du sol. J.Appl. Genet. 2020 , 61 , 249-263. [ Google Scholar ] [ CrossRef ]
Parada, AE ; Needham, DM; Fuhrman, JA Chaque base compte : évaluation des amorces d’ARNr de petites sous-unités pour les microbiomes marins avec des communautés simulées, des séries chronologiques et des échantillons mondiaux de terrain. Environ. Microbiol. 2016 , 18 , 1403-1414. [ Google Scholar ] [ CrossRef ]
Bokulich, NA; Mills, DA Une sélection améliorée d’amorces transcrites spécifiques aux espaceurs internes permet un profilage quantitatif à ultra-haut débit des communautés fongiques. Appl. Environ. Microbiol. 2013 , 79 , 2519-2526. [ Google Scholar ] [ CrossRef ]
Comeau, AM; Li, WKW; Tremblay, J.-É.; Carmack, CE; Lovejoy, C. Structure de la communauté microbienne de l’océan Arctique avant et après le minimum record de glace de mer de 2007. PLoS ONE 2011 , 6 , e27492. [ Google Scholar ] [ CrossRef ]
Emerson, JB ; Thomas, Colombie-Britannique ; Alvarez, W. ; Banfield, JF Analyse métagénomique d’une communauté microbienne souterraine à haute teneur en dioxyde de carbone peuplée de chimiolithoautotrophes, de bactéries et d’archées provenant de phylums candidats. Environ. Microbiol. 2016 , 18 , 1686-1703. [ Google Scholar ] [ CrossRef ]
Castellano-Hinojosa, A. ; González-López, J. ; Vallejo, A. ; Bedmar, EJ Effet des inhibiteurs d’uréase et de nitrification sur la volatilisation de l’ammoniac et l’abondance des gènes du cycle N dans un sol agricole. J. Plante Nutr. Science du sol. 2020 , 183 , 99-109. [ Google Scholar ] [ CrossRef ]
Morales, MOI ; Allegrini, M. ; Basualdo, J. ; Villamil, Manitoba ; Zabaloy, conception MC Primer pour évaluer la dégradation bactérienne du glyphosate et d’autres phosphonates. J. Microbiol. Méthodes 2020 , 169 , 105814. [ Google Scholar ] [ CrossRef ]
Kanehisa, M. ; Furumichi, M. ; Sato, Y. ; Kawashima, M. ; Ishiguro-Watanabe, M. KEGG pour l’analyse taxonomique des voies et des génomes. Acides nucléiques Res. 2023 , 51 , D587-D592. [ Google Scholar ] [ CrossRef ]
Nilsson, RH; Kristiansson, E. ; Ryberg, M. ; Hallenberg, N. ; Larsson, KH Variabilité intraspécifique des ITS dans le règne des champignons telle qu’exprimée dans les bases de données de séquences internationales et ses implications pour l’identification des espèces moléculaires. Évol. Bioinformer. En ligne 2008 , 4 , 193-201. [ Google Scholar ] [ CrossRef ]
Kepler, RM; Epp Schmidt, DJ ; Yarwood, SA ; Cavigelli, MA; Reddy, KN ; Duc, DONC ; Bradley, Californie ; Williams, MM; Acheteur, JS ; Maul, JE Communautés microbiennes du sol dans divers agroécosystèmes exposés à l’herbicide glyphosate. Appl. Environ. Microbiol. 2020 , 86 , e01744-19. [ Google Scholar ] [ CrossRef ]
Dennis, PG ; Kukulies, T. ; Forstner, C. ; Orton, TG; Pattison, AB Les effets du glyphosate, du glufosinate, du paraquat et du paraquat-diquat sur l’activité microbienne du sol et sur la diversité bactérienne, archéenne et nématode. Sci. Rep. 2018 , 8 , 2119. [ Google Scholar ] [ CrossRef ]
Tisserand, MA ; Krutz, LJ; Zablotowicz, RM; Reddy, KN Effets du glyphosate sur les communautés microbiennes du sol et sa minéralisation dans un sol du Mississippi. Gestion antiparasitaire. Sci. 2007 , 63 , 388-393. [ Google Scholar ] [ CrossRef ]
Guijarro, KH; Aparicio, V. ; De Gerónimo, E. ; Castellote, M. ; Figuerola, EL; Costa, JL; Erijman, L. Communautés microbiennes du sol et dégradation du glyphosate dans des sols ayant des antécédents d’application d’herbicides différents. Sci. Environ. Total. 2018 , 634 , 974-982. [ Google Scholar ] [ CrossRef ]
Lupwayi, Nouvelle-Zélande ; Larney, FJ; Blackshaw, RE; Kanashiro, DA; Pearson, DC ; Petri, RM Le pyroséquençage révèle les profils des communautés bactériennes du sol après 12 ans de gestion de conservation sur les rotations de cultures irriguées. Appl. Ecol du sol. 2017 , 121 , 65-73. [ Google Scholar ] [ CrossRef ]
Schmidt, R. ; Graver, K. ; Bossange, AV; Mitchell, J. ; Scow, K. Utilisation à long terme de cultures de couverture et stratégies de vie de la communauté microbienne du sol sans labour dans les sols agricoles. PLoS ONE 2018 , 13 , e0192953. [ Google Scholar ] [ CrossRef ]
Giusti, B. ; Hogue, R. ; Jeanne, T. ; Lucotte, M. Impacts du blé d’hiver et des cultures de couverture sur la diversité microbienne du sol dans un système de culture sans labour maïs-soja au Québec (Canada). Agrosyste. Géosci. Environ. 2023 , 6 , e20349. [ Google Scholar ] [ CrossRef ]
Samson-Brais, É.; Lucotte, M. ; Moingt, M. ; Tremblay, G. ; Paquet, S. Teneurs en glyphosate et en acide aminométhylphosphonique dans les sols de grandes cultures selon diverses pratiques de lutte contre les mauvaises herbes. Agrosyste. Géosci. Environ. 2022 , 5 , e20273. [ Google Scholar ] [ CrossRef ]
Alferness, PL; Iwata, Y. Détermination du glyphosate et de l’acide (aminométhyl)phosphonique dans les matrices de sol, végétales et animales et dans l’eau par chromatographie en phase gazeuse capillaire avec détection sélective de masse. J. Agric. Chimie alimentaire. 1994 , 42 , 2751-2759. [ Google Scholar ] [ CrossRef ]
Conseil des Productions Végétales du Québec. Méthode MA-2 ; Ministère de l’Agriculture, des Pêcheries et de l’Alimentation (Québec) : Québec, QC, Canada, 1993. [ Google Scholar ]
Centre d’Expertise en Analyse Environnementale du Québec. Détermination du pH : Méthode Électrométrique ; Ministère de l’Environnement, de la Lutte contre les changements climatiques, de la Faune et des Parcs : Québec, QC, Canada, 2014; Tome 3. [ Google Scholar ]
Cordonnier, HE; McLean, EO; Pratt, PF Méthodes tampons pour déterminer les besoins en chaux des sols contenant des quantités appréciables d’aluminium extractible. Science du sol. Soc. Suis. J. 1961 , 25 , 274-277. [ Google Scholar ] [ CrossRef ]
Mehlich, A. Extracteur d’analyse de sol Mehlich 3 : Une modification de l’extracteur Mehlich 2. Commun. Science du sol. Plante Anale. 1984 , 15 , 1409-1416. [ Google Scholar ] [ CrossRef ]
Culman, S. ; Mann, M. ; Brown, C. Calcul de la capacité d’échange de cations, de la saturation en base et de la saturation en calcium. 2019. Disponible en ligne : https://ohioline.osu.edu/factsheet/anr-81 (consulté le 19 février 2024).
CRAAQ. Guide de référence sur la fertilisation , 2e éd.; CRAAQ : Québec, QC, Canada, 2010. (En français) [ Google Scholar ]
Apprill, A. ; McNally, S. ; Parsons, R. ; Weber, L. Une révision mineure de l’amorce du gène SSU rRNA 806R de la région V4 augmente considérablement la détection du bactérioplancton SAR11. Aquat. Microbe. Écol. 2015 , 75 , 129-137. [ Google Scholar ] [ CrossRef ]
Jeanne, T. ; Parent, S.-É.; Hogue, R. Utilisation d’un indice d’équilibre des espèces bactériennes du sol pour estimer la productivité des cultures de pommes de terre. PLoS ONE 2019 , 14 , e0214089. [ Google Scholar ] [ CrossRef ] [ PubMed ]
Bolyen, E. ; Rideout, JR ; Dillon, M. ; Bokulich, NA; Abnet, CC; Al-Ghalith, Géorgie ; Alexandre, H. ; Alm, EJ; Arumugam, M. ; Asnicar, F. ; et coll. Science des données du microbiome reproductible, interactive, évolutive et extensible à l’aide de QIIME 2. Nat. Biotechnologie. 2019 , 37 , 852-857. [ Google Scholar ] [ CrossRef ]
Quast, C. ; Pruesse, E. ; Yilmaz, P. ; Gerken, J. ; Schweer, T. ; Yarza, P. ; Péplies, J. ; Glöckner, FO Le projet de base de données de gènes d’ARN ribosomal SILVA : traitement des données amélioré et outils Web. Acides nucléiques Res. 2013 , 41 , D590-D596. [ Google Scholar ] [ CrossRef ] [ PubMed ]
Kõljalg, U. ; Nilsson, RH; Abarenkov, K. ; Tedersoo, L. ; Taylor, AFS; Bahram, M. ; Bates, ST ; Bruns, TD; Bengtsson-Palme, J. ; Callaghan, TM; et coll. Vers un paradigme unifié pour l’identification des champignons basée sur des séquences. Mol. Écol. 2013 , 22 , 5271-5277. [ Google Scholar ] [ CrossRef ] [ PubMed ]
Callahan, BJ; McMurdie, PJ ; Holmes, SP Les variantes de séquence exactes devraient remplacer les unités taxonomiques opérationnelles dans l’analyse des données sur les gènes marqueurs. ISME J. 2017 , 11 , 2639-2643. [ Google Scholar ] [ CrossRef ]
Overbeek, W. ; Jeanne, T. ; Hogue, R. ; Smith, DL Effets des consortiums microbiens, appliqués comme revêtement d’engrais, sur les communautés microbiennes du sol et de la rhizosphère et sur le rendement des pommes de terre. Devant. Agron. 2021 , 3 , 714700. [ Google Scholar ] [ CrossRef ]
Jeanne, T. ; D’Astous-Pagé, J.; Hogue, R. Variabilité spatiale, temporelle et technique de la diversité des procaryotes et des champignons dans les sols agricoles. Devant. Science du sol. 2022 , 2 , 945888. [ Google Scholar ] [ CrossRef ]
Douglas, directeur général ; Maffei, VJ; Zaneveld, JR; Yurgel, SN; Brun, JR ; Taylor, CM ; Huttenhower, C. ; Langille, MGI PICRUSt2 pour la prédiction des fonctions du métagénome. Nat. Biotechnologie. 2020 , 38 , 685-688. [ Google Scholar ] [ CrossRef ]
Caspi, R. ; Billington, R. ; Ferrer, L. ; Foerster, H. ; Fulcher, Californie ; Keseler, IM; Kothari, A. ; Krummenacker, M. ; Latendresse, M. ; Mueller, LA ; et coll. La base de données MetaCyc des voies métaboliques et des enzymes et la collection BioCyc de bases de données de voies/génomes. Acides nucléiques Res. 2016 , 44 , D471-D480. [ Google Scholar ] [ CrossRef ]
Hernández Guijarro, K. ; De Gerónimo, E. ; Erijman, L. Potentiel de biodégradation du glyphosate dans le sol basé sur le gène de la glycine oxydase (thiO) de Bradyrhizobium. Curr. Microbiol. 2021 , 78 , 1991-2000. [ Google Scholar ] [ CrossRef ]
Hove-Jensen, B. ; Zechel, DL; Jochimsen, B. Utilisation du glyphosate comme phosphate Source : Biochimie et génétique de la lyase carbone-phosphore bactérienne. Microbiol. Mol. Biol. Rév.2014 , 78 , 176-197 . [ Google Scholar ] [ CrossRef ] [ PubMed ]
Équipe de base R. R : Un langage et un environnement pour le calcul statistique ; R Foundation for Statistical Computing : Vienne, Autriche, 2020. [ Google Scholar ]
McMurdie, PJ ; Holmes, S. Ne gaspillez pas, ne voulez pas : pourquoi la raréfaction des données sur le microbiome est inacceptable. Calcul PLoS. Biol. 2014 , 10 , e1003531. [ Google Scholar ] [ CrossRef ] [ PubMed ]
McMurdie, PJ ; Holmes, S. phyloseq : Un package R pour une analyse interactive reproductible et des graphiques des données de recensement du microbiome. PLoS ONE 2013 , 8 , e61217. [ Google Scholar ] [ CrossRef ] [ PubMed ]
Oksanen, J. ; Blanchet, G. ; Amicalement, M. ; Kindt, R. ; Legendre, P. ; McGlinn, D. ; Minchin, P. ; O’Hara, R. ; Simpson, G. ; Solymos, P. ; et coll. Végétalien : Forfait Écologie Communautaire. 2019. Disponible en ligne : http://CRAN.Rproject.org/package=vegan (consulté le 14 février 2024).
Daughton Christian, G. Gestion du berceau au berceau des médicaments pour minimiser leur élimination dans l’environnement tout en favorisant la santé humaine. I. Justification et pistes vers une pharmacie verte. Environ. Perspective de santé. 2003 , 111 , 757-774. [ Google Scholar ] [ CrossRef ] [ PubMed ]
Primost, J. ; Marin, D. ; Aparicio, V. ; Costa, J. ; Carriquiriborde, P. Glyphosate et AMPA, polluants « pseudo-persistants » dans le cadre de pratiques de gestion agricole réelles dans l’agroécosystème mésopotamique de la Pampa, Argentine. Environ. Polluer. 2017 , 229 , 771-779. [ Google Scholar ] [ CrossRef ] [ PubMed ]
Fuchs, B. ; Saikkonen, K. ; Hélander, M. ; Tian, Y. ; Yang, B. ; Engström, MT ; Salminen, J.-P. ; Muola, A. Héritage des produits agrochimiques dans l’économie alimentaire circulaire : les herbicides à base de glyphosate introduits via le fumier affectent le rendement et la biochimie des plantes cultivées vivaces au cours de l’année suivante. Chemosphere 2022 , 308 , 136366. [ Google Scholar ] [ CrossRef ] [ PubMed ]
Giard, F. ; Lucotte, M. ; Moingt, M. ; Gaspar, A. Teneur en glyphosate et en aminométhyphosphonique (AMPA) dans les sols des grandes cultures brésiliennes. Agron. Sci. Biotechnologie. 2022 , 8 , 1–18. [ Google Scholar ] [ CrossRef ]
Silva, V. ; Montanarella, L. ; Jones, A. ; Fernández-Ugalde, O. ; Mol, HGJ; Ritsema, CJ; Geissen, V. Distribution du glyphosate et de l’acide aminométhylphosphonique (AMPA) dans les couches arables agricoles de l’Union européenne. Sci. Environ. Total. 2018 , 621 , 1352-1359. [ Google Scholar ] [ CrossRef ]
Mamy, L. ; Barriuso, E. ; Gabrielle, B. Devenir du glyphosate dans les sols lorsqu’il arrive dans les résidus végétaux. Chemosphère 2016 , 154 , 425-433. [ Google Scholar ] [ CrossRef ] [ PubMed ]
Mencaroni, M. ; Cardinali, A. ; Costa, L. ; Morari, F. ; Salandin, P. ; Zanin, G. ; Dal Ferro, N. Glyphosate et AMPA ont une faible mobilité à travers différents profils de sol de la zone de production de vin prosecco : une étude de surveillance dans le nord-est de l’Italie. Devant. Environ. Sci. 2022 , 10 , 971931. [ Google Scholar ] [ CrossRef ]
Vereecken, H. Mobilité et lixiviation du glyphosate : une revue. Gestion antiparasitaire. Sci. 2005 , 61 , 1139-1151. [ Google Scholar ] [ CrossRef ] [ PubMed ]
Cassigneul, A. ; Benoit, P. ; Bergheaud, V. ; Dumény, V. ; Etiévant, V. ; Goubard, Y. ; Maylin, A. ; Justes, E. ; Alletto, L. Devenir du glyphosate et de ses produits de dégradation dans les résidus de cultures de couverture et le sol sous-jacent : une étude en laboratoire. Sci. Environ. Total. 2016 , 545-546 , 582-590. [ Google Scholar ] [ CrossRef ] [ PubMed ]
Aslam, S. ; Iqbal, A. ; Lafolie, F. ; Recous, S. ; Benoit, P. ; Garnier, P. Le paillis de résidus végétaux à la surface du sol impacte le lessivage et la persistance des pesticides : Une étude de modélisation à partir de colonnes de sol. J. Contam. Hydrol. 2018 , 214 , 54-64. [ Google Scholar ] [ CrossRef ]
Aslam, S. ; Jing, Y. ; Nowak, KM Devenir du glyphosate et de ses produits de dégradation AMPA, glycine et sarcosine dans un sol agricole : implications pour l’évaluation des risques environnementaux. J. Hazard. Maître. 2023 , 447 , 130847. [ Google Scholar ] [ CrossRef ] [ PubMed ]
Samson-Brais, É.; Lucotte, M. ; Moingt, M. ; Tremblay, G. ; Paquet, S. Impact des pratiques de gestion des mauvaises herbes sur l’activité biologique des sols de grandes cultures de maïs et de soja au Québec (Canada). Peut. J. Sol Sci. 2020 , 101 , 12-21. [ Google Scholar ] [ CrossRef ]
Voie, M. ; Lorenz, N. ; Saxena, J. ; Ramsier, C. ; Dick, R. L’effet du glyphosate sur l’activité microbienne du sol, la structure de la communauté microbienne et le potassium du sol. Pédobiologie 2012 , 55 , 335-342. [ Google Scholar ] [ CrossRef ]
McConnell, JS; Hossner, LR Isothermes d’adsorption dépendant du pH du glyphosate. J. Agric. Chimie alimentaire. 1985 , 33 , 1075-1078. [ Google Scholar ] [ CrossRef ]
de Jonge, H. ; Wollesen de Jonge, L. Influence du pH et de la composition de la solution sur la sorption du glyphosate et du prochloraz dans un sol limoneux sableux. Chemosphère 1999 , 39 , 753-763. [ Google Scholar ] [ CrossRef ]
Weber, JB ; Wilkerson, GG; Reinhardt, CF Calcul des coefficients de sorption des pesticides (Kd) à l’aide de propriétés de sol sélectionnées. Chemosphère 2004 , 55 , 157-166. [ Google Scholar ] [ CrossRef ]
Kumari, KGID ; Moldrup, P. ; Paradelo, M. ; Elsgaard, L. ; de Jonge, LW Les propriétés du sol contrôlent la sorption du glyphosate dans les sols modifiés avec du biochar de bois de bouleau. Pollution de l’eau, de l’air et du sol. 2016 , 227 , 174. [ Google Scholar ] [ CrossRef ]
De Gerónimo, E. ; Aparicio, VC Les changements dans le pH du sol et l’ajout de phosphate inorganique affectent l’adsorption du glyphosate dans le sol agricole. EUR. J. Sol Sci. 2022 , 73 , e13188. [ Google Scholar ] [ CrossRef ]
Sidoli, P. ; Baran, N. ; Angulo-Jaramillo, R. Adsorption du glyphosate et de l’AMPA dans les sols : expériences de laboratoire et règles de pédotransfert. Environ. Sci. Polluer. Rés. 2016 , 23 , 5733-5742. [ Google Scholar ] [ CrossRef ] [ PubMed ]
Li, Y. ; Chanson, D. ; Liang, S. ; Dan, P. ; Qin, X. ; Liao, Y. ; Siddique, KHM Effet du non-labour sur la diversité des communautés bactériennes et fongiques du sol : une méta-analyse. Travail du sol Res. 2020 , 204 , 104721. [ Google Scholar ] [ CrossRef ]
Soleil, R. ; Li, W. ; Dong, W. ; Tian, Y. ; Hu, C. ; Liu, B. Le travail du sol modifie la répartition verticale des communautés bactériennes et fongiques du sol. Devant. Microbiol. 2018 , 9 , 699. [ Google Scholar ] [ CrossRef ] [ PubMed ]
Gao, M. ; Li, H. ; Li, M. Effet du système sans labour sur la structure de la communauté fongique du sol des terres cultivées à Mollisol : une étude de cas. Devant. Microbiol. 2022 , 13 , 847691. [ Google Scholar ] [ CrossRef ] [ PubMed ]
Lu, X. ; Lu, X. ; Liao, Y. Effet du traitement du sol sur la diversité de la teneur en carbone des champignons mycorhiziens arbusculaires du sol et des agrégats du sol. Devant. Microbiol. 2018 , 9 , 410634. [ Google Scholar ] [ CrossRef ]
Liu, W. ; Maman, K. ; Wang, X. ; Wang, Z. ; Negrete-Yankelevich, S. Effets du sans labour et des engrais organiques d’origine biologique sur les communautés fongiques mycorhiziennes arbusculaires du sol dans les champs de blé d’hiver. Appl. Ecol du sol. 2022 , 178 , 104564. [ Google Scholar ] [ CrossRef ]
Zhang, W.-w.; Wang, C. ; Xue, R. ; Wang, L.-j. Effets de la salinité sur la communauté microbienne du sol et la fertilité du sol. J. Intégré. Agricole. 2019 , 18 , 1360-1368. [ Google Scholar ] [ CrossRef ]
Rath, KM; Rousk, J. Effets du sel sur la communauté des décomposeurs microbiens du sol et leur rôle dans le cycle du carbone organique : une revue. Biol du sol. Biochimie. 2015 , 81 , 108-123. [ Google Scholar ] [ CrossRef ]
Obayomi, O. ; Seyoum, MM; Ghazaryan, L. ; Tebbé, CC ; Murase, J. ; Bernstein, N. ; Gillor, O. La texture et les propriétés du sol plutôt que le type d’eau d’irrigation façonnent la diversité et la composition des communautés microbiennes du sol. Appl. Ecol du sol. 2021 , 161 , 103834. [ Google Scholar ] [ CrossRef ]
Xia, Q. ; Rufty, T. ; Shi, W. Diversité et composition microbiennes du sol : liens avec la texture du sol et les propriétés associées. Biol du sol. Biochimie. 2020 , 149 , 107953. [ Google Scholar ] [ CrossRef ]
Mann, C. ; Lynch, D. ; Fillmore, S. ; Mills, A. Relations entre la gestion des champs, la santé des sols et la composition de la communauté microbienne. Appl. Ecol du sol. 2019 , 144 , 12-21. [ Google Scholar ] [ CrossRef ]
Venter, ZS ; Jacobs, K. ; Hawkins, H.-J. L’impact de la rotation des cultures sur la diversité microbienne du sol : une méta-analyse. Pédobiologie 2016 , 59 , 215-223. [ Google Scholar ] [ CrossRef ]
Guo, Z. ; Wan, S. ; Hua, K. ; Yin, Y. ; Chu, H. ; Wang, D. ; Guo, X. Le régime de fertilisation a un effet plus important sur la structure de la communauté microbienne du sol que la rotation des cultures et le stade de croissance dans un agroécosystème. Appl. Ecol du sol. 2020 , 149 , 103510. [ Google Scholar ] [ CrossRef ]
Kracmarova, M. ; Uhlik, O. ; Strejcek, M. ; Szakova, J. ; Cerny, J. ; Balik, J. ; Tlustos, P. ; Kohout, P. ; Demnerova, K. ; Stiborova, H. Communautés microbiennes du sol après 20 ans de pratiques de fertilisation et de rotation des cultures en République tchèque. Environ. Microbiome 2022 , 17 , 13. [ Google Scholar ] [ CrossRef ] [ PubMed ]
Ville, JR ; Dumonceaux, T. ; Tidemann, B. ; Helgason, BL La rotation des cultures influence de manière significative la composition du sol, de la rhizosphère et du microbiote racinaire du canola ( Brassica napus L.). Environ. Microbiome 2023 , 18 , 40. [ Google Scholar ] [ CrossRef ] [ PubMed ]
Hart, MM; Powell, JR ; Gulden, RH; Dunfield, KE ; Peter Pauls, K. ; Swanton, CJ; Klironomos, JN; Antunes, PM ; Koch, UN M; Trevors, JT Séparer l’effet de la culture de l’herbicide sur les communautés microbiennes du sol dans le maïs résistant au glyphosate. Pédobiologie 2009 , 52 , 253-262. [ Google Scholar ] [ CrossRef ]
Daniel, CS; Yin, C. ; Hulbert, S. ; Burke, moi; Paulitz, T. Les impacts de l’utilisation répétée du glyphosate sur les bactéries associées au blé sont faibles et dépendent de l’historique d’utilisation du glyphosate. Appl. Environ. Microbiol. 2017 , 83 , e01354-17. [ Google Scholar ] [ CrossRef ]
Avis de non-responsabilité/Note de l’éditeur : Les déclarations, opinions et données contenues dans toutes les publications sont uniquement celles du ou des auteurs et contributeurs individuels et non de MDPI et/ou du ou des éditeurs. MDPI et/ou le(s) éditeur(s) déclinent toute responsabilité pour tout préjudice corporel ou matériel résultant des idées, méthodes, instructions ou produits mentionnés dans le contenu.
James J. Hoorman, cultures de couverture et qualité de l’eau, éducateur en vulgarisation, vulgarisation de l’Ohio State UniversityRafiq Islam, spécialiste des sols et de l’eau, Ohio State University Extension, South Centers at Piketon
Les micro-organismes du sol existent en grand nombre dans le sol tant qu’il existe une source de carbone pour l’énergie. Il existe un grand nombre de bactéries dans le sol, mais en raison de leur petite taille, leur biomasse est plus faible. Les actinomycètes sont 10 fois plus petits en nombre, mais sont plus grands en taille et ont donc une biomasse similaire à celle des bactéries. Les populations de champignons sont plus petites mais dominent la biomasse du sol lorsque le sol n’est pas perturbé. Les bactéries, les actinomycètes et les protozoaires sont robustes et peuvent tolérer davantage de perturbations du sol que les populations fongiques. Ils dominent donc dans les sols labourés, tandis que les populations fongiques et nématodes ont tendance à dominer dans les sols labourés ou sans labour.
Il y a plus de microbes dans une cuillère à café de terre qu’il n’y a d’habitants sur terre. Les sols contiennent environ 8 à 15 tonnes de bactéries, champignons, protozoaires, nématodes, vers de terre et arthropodes. Voir les fiches d’information sur les rôles des bactéries, des champignons, des protozoaires et des nématodes du sol.
Tableau 1 : Nombre relatif et biomasse des espèces microbiennes à une profondeur de sol de 0 à 6 pouces (0 à 15 cm)
Microorganismes
Nombre/g de sol
Biomasse (g/m 2 )
Bactéries
10 8 –10 9
40 à 500
Actinomycètes
10 7 –10 8
40 à 500
Champignons
10 5 –10 6
100-1500
Algues
10 4 –10 5
1 à 50
Protozoaires
10 3 –10 4
Varie
Nématodes
10 2 –10 3
Varie
Décomposition microbienne de la matière organique du sol
La décomposition de la matière organique remplit deux fonctions pour les micro-organismes : fournir de l’énergie pour la croissance et fournir du carbone pour la formation de nouvelles cellules. La matière organique du sol (MOS) est composée des fractions « vivantes » (micro-organismes), « mortes » (résidus frais) et « très mortes » (humus). Le « très mort » ou humus est la fraction de MOS à long terme, vieille de plusieurs milliers d’années et résistante à la décomposition. La matière organique du sol contient deux composants appelés MOS active (35 %) et passive (65 %). La MOS active est composée de matières végétales ou animales fraîches « vivantes » et « mortes » qui servent de nourriture aux microbes et sont composées de sucres et de protéines faciles à digérer. La SOM passive est résistante à la décomposition par les microbes et est plus riche en lignine.
Les microbes ont besoin d’un apport régulier de MOS active dans le sol pour survivre dans le sol. Les sols non labourés à long terme contiennent des niveaux de microbes nettement plus élevés, plus de carbone actif, plus de MOS et plus de carbone stocké que les sols labourés conventionnels. La majorité des microbes présents dans le sol existent dans des conditions de famine et ont donc tendance à être dans un état dormant, en particulier dans les sols labourés.
Les résidus végétaux morts et les éléments nutritifs des plantes deviennent la nourriture des microbes présents dans le sol. La matière organique du sol (MOS) correspond essentiellement à toutes les substances organiques (tout ce qui contient du carbone) présentes dans le sol, vivantes et mortes. La MOS comprend les plantes, les algues bleu-vert, les micro-organismes (bactéries, champignons, protozoaires, nématodes, coléoptères, collemboles, etc.) et la matière organique fraîche et en décomposition provenant des plantes, des animaux et des micro-organismes.
La matière organique du sol peut être décomposée en ses éléments constitutifs. Cent grammes (g) ou 100 livres (lbs) de matière végétale morte produisent environ 60 à 80 g (lbs) de dioxyde de carbone, qui est rejeté dans l’atmosphère. Les 20 à 40 g (lb) d’énergie et de nutriments restants sont décomposés et transformés en environ 3 à 8 g (lb) de micro-organismes (les vivants), 3 à 8 g (lb) de composés non humiques (les morts), et 10 à 30 g (lb) d’humus (la matière très morte, résistante à la décomposition). La structure moléculaire de la MOS est principalement constituée de carbone et d’oxygène avec un peu d’hydrogène et d’azote et de petites quantités de phosphore et de soufre. La matière organique du sol est un sous-produit des cycles du carbone et de l’azote.
Figure 1. Diagramme du Dr Rafiq Islam.
Nutriments de la matière organique du sol
Les éléments nutritifs du sol ont une valeur actuelle de 680 $ pour chaque 1 pour cent de MOS ou de 68 $ par tonne de MOS sur la base des valeurs économiques des engrais commerciaux (voir tableau 2). La MOS est composée principalement de carbone, mais associée au carbone sont de grandes quantités d’azote et de soufre provenant des protéines, du phosphore et du potassium. Le SOM doit être considéré comme un investissement dans un certificat de dépôt (CD). Les sols biologiquement actifs et contenant de plus grandes quantités de carbone actif recyclent et libèrent plus de nutriments pour la croissance des plantes que les sols biologiquement inactifs et contiennent moins de matière organique active. Dans des conditions de culture sans labour, de petites quantités de nutriments sont libérées chaque année (comme les intérêts sur un CD) pour fournir des nutriments lentement et efficacement aux racines des plantes. Cependant, avec le travail du sol, de grandes quantités de nutriments peuvent être libérées puisque la MOS est consommée et détruite par les microbes. Étant donné que les niveaux de MOS mettent du temps à s’accumuler, la capacité de stockage des nutriments est diminuée et les excédents de nutriments libérés sont souvent lessivés dans les eaux de surface. La SOM est un réservoir de nombreux nutriments végétaux.
Considérez les trois scénarios suivants. Les sols renouvellent généralement 1 à 3 pour cent de leur azote stocké dans la MOS. Les sols labourés ou malsains libèrent un pourcentage d’azote plus faible en raison d’une activité microbienne plus faible. Un sol labouré avec 2 pour cent de MOS (2 000 livres de N) peut libérer 1 pour cent de N ou 20 livres de N par an. Un sol plus biologiquement actif et contenant 4 pour cent de MOS (4 000 lb de N) peut libérer 1,5 pour cent de N ou 60 lb de N, tandis qu’un sol à 6 pour cent de MOS (6 000 lb de N) peut libérer 2 pour cent de N ou 120 lb de N. Dans les sols, les excédents de nutriments libérés sont souvent perdus et les réserves de carbone sont épuisées, ce qui réduit le stockage futur des nutriments. Les agriculteurs constatent souvent cela lorsqu’ils labourent un sol vierge, un vieux pâturage ou une clôture. Pendant plusieurs années, les cultures sur le sol nouvellement labouré pousseront mieux que les sols environnants, mais avec le temps, le sol sera épuisé en carbone et le sol nouvellement labouré deviendra moins fertile car le carbone est oxydé sous forme de dioxyde de carbone et perdu dans l’atmosphère. . Le travail du sol entraîne l’oxydation et la destruction du carbone dans le sol en augmentant les niveaux d’oxygène du sol, favorisant ainsi l’expansion des populations de bactéries et la consommation de carbone actif dans le sol.
Tableau 2 : Valeur de la matière organique du sol
Hypothèses : 2 000 000 livres de sol dans les 6 premiers pouces
Nutriments
1% de matière organique = 20 000# 50%Carbone, rapport C:N = 10:1
Effets du climat, de la température et du pH sur la MOS
La MOS est affectée par le climat et la température. Les populations microbiennes doublent à chaque changement de température de 10 degrés Fahrenheit. Si nous comparons les tropiques aux régions arctiques plus froides, nous constatons que la majeure partie du carbone est retenue dans les arbres et la végétation aérienne. Sous les tropiques, la couche arable contient très peu de MOS car les températures élevées et l’humidité décomposent rapidement la MOS. En se déplaçant vers le nord ou le sud de l’équateur, la MOS augmente dans le sol. La toundra près du cercle polaire arctique contient une grande quantité de MOS en raison des températures froides. Les températures glaciales modifient le sol de sorte qu’une plus grande quantité de MOS est décomposée que dans les sols non sujets au gel.
L’humidité, le pH, la profondeur du sol et la taille des particules affectent la décomposition de la MOS. Les régions chaudes et humides stockent moins de carbone organique dans le sol que les régions sèches et froides en raison d’une décomposition microbienne accrue. Le taux de décomposition de la MOS augmente lorsque le sol est exposé à des cycles de séchage et d’humidification par rapport aux sols continuellement humides ou secs. Toutes choses étant égales par ailleurs, les sols dont le pH est neutre à légèrement alcalin décomposent la MOS plus rapidement que les sols acides ; par conséquent, le chaulage du sol améliore la décomposition de la MOS et l’évolution du dioxyde de carbone. La décomposition est également plus importante près de la surface du sol, là où se trouvent la plus forte concentration de résidus végétaux. À de plus grandes profondeurs, la décomposition de la MOS est moindre, ce qui correspond à une baisse des niveaux de carbone organique due à la diminution des résidus végétaux. Les petites particules sont plus facilement dégradées par les microbes du sol que les grosses particules, car la surface globale est plus grande avec les petites particules, de sorte que les microbes peuvent attaquer les résidus.
Une différence dans la formation des sols se produit également d’est en ouest à travers les États-Unis. À l’est, les forêts de feuillus prédominaient et les racines pivotantes des arbres étaient riches en lignine, et les arbres à feuilles caduques laissaient de grandes quantités de feuilles mortes à la surface du sol. Les racines des feuillus ne se renouvellent pas rapidement, de sorte que les niveaux de matière organique dans le sous-sol sont assez faibles. Dans les sols forestiers, la majeure partie de la MOS est distribuée dans les quelques centimètres supérieurs. À mesure que vous vous déplacez vers l’ouest, de hautes prairies dominent le paysage et la couche arable se forme à partir de systèmes racinaires profonds et fibreux. Cinquante pour cent des racines d’une herbe meurent et sont remplacées chaque année et les racines de l’herbe sont riches en sucres et en protéines (matière organique active plus élevée) et plus faibles en lignine. Ainsi, les sols formés sous les prairies à herbes hautes sont riches en MOS dans tout le profil pédologique. Ces sols de premier choix sont très productifs car ils ont un pourcentage plus élevé de MOS (en particulier de charbon actif), retiennent plus de nutriments, contiennent plus de microbes et ont une meilleure structure du sol en raison de populations fongiques plus importantes.
Rapport carbone/azote
La dégradation des résidus organiques par les microbes dépend du rapport carbone/azote (C:N). Les microbes présents dans le rumen d’une vache, dans un tas de compost et dans le sol dépendent du rapport C:N pour décomposer les résidus organiques (à base de carbone). Considérez deux sources d’alimentation distinctes, un jeune plant de luzerne tendre et de la paille d’avoine ou de blé. Une jeune plante de luzerne contient plus de protéines brutes, d’acides aminés et de sucres dans la tige, elle est donc facilement digérée par les microbes, que ce soit dans le rumen d’une vache, dans un tas de compost ou dans le sol. La jeune luzerne a une teneur élevée en azote provenant des protéines (les acides aminés et les protéines sont riches en azote et en soufre), elle a donc un rapport carbone/azote plus faible (moins de carbone, plus d’azote). Cependant, la paille d’avoine et de blé (ou le foin plus âgé) contient plus de lignine (qui résiste à la décomposition microbienne), moins de protéines brutes, moins de sucres dans la tige et un rapport C:N plus élevé. La paille est décomposée par les microbes, mais il faut plus de temps et d’azote pour décomposer cette source riche en carbone.
Une faible teneur en azote ou un rapport C:N élevé est associé à une lente décomposition de la MOS. Les plantes immatures ou jeunes ont une teneur en azote plus élevée, des rapports C:N plus faibles et une décomposition plus rapide de la SOM. Pour un bon compostage, un rapport C:N inférieur à 20 permet aux matières organiques de se décomposer rapidement (4 à 8 semaines) tandis qu’un rapport C:N supérieur à 20 nécessite un supplément d’azote et ralentit la décomposition. Ainsi, si nous ajoutons au sol un matériau à haute teneur en carbone et à faible teneur en N, les microbes capteront l’azote du sol. Finalement, l’azote du sol est libéré mais à court terme, l’azote est retenu. Le facteur de conversion pour convertir l’azote en protéines brutes est de 16,7, ce qui explique pourquoi il est si important d’avoir un rapport C:N inférieur à 20.
Le rapport C:N de la plupart des sols est d’environ 10:1, ce qui indique que l’azote est disponible pour la plante. Le rapport C:N de la plupart des résidus végétaux a tendance à diminuer avec le temps, à mesure que la MOS se désintègre. Cela résulte de la perte gazeuse de dioxyde de carbone. Par conséquent, le pourcentage d’azote dans la MOS résiduelle augmente à mesure que la décomposition progresse. Le rapport C:N de 10:1 de la plupart des sols reflète une valeur d’équilibre associée à la plupart des microbes du sol (bactéries 3:1 à 10:1, champignon 10:1 rapport C:N).
Les bactéries sont les premiers microbes à digérer les nouveaux résidus organiques végétaux et animaux présents dans le sol. Les bactéries peuvent généralement se reproduire en 30 minutes et ont une teneur élevée en N dans leurs cellules (3 à 10 atomes de carbone pour 1 atome d’azote ou 10 à 30 pour cent d’azote). Dans de bonnes conditions de chaleur, d’humidité et de nourriture, ils peuvent se reproduire très rapidement. Les bactéries sont généralement moins efficaces pour convertir le carbone organique en nouvelles cellules. Les bactéries aérobies assimilent environ 5 à 10 pour cent du carbone tandis que les bactéries anaérobies n’en assimilent que 2 à 5 pour cent, laissant derrière elles de nombreux déchets composés de carbone et utilisant de manière inefficace l’énergie stockée dans la MOS.
Figure 2. Graphique du N relatif disponible avec la durée de décomposition.
Luzerne, faible rapport C:N, C:N = 13:1
Paille d’avoine, rapport C:N élevé, C:N = 80:1
Les champignons libèrent généralement moins de dioxyde de carbone dans l’atmosphère et sont plus efficaces pour convertir le carbone pour former de nouvelles cellules. Le champignon capte généralement plus d’énergie de la MOS à mesure qu’il la décompose, assimilant 40 à 55 pour cent du carbone. La plupart des champignons consomment de la matière organique plus riche en cellulose et en lignine, dont la décomposition est plus lente et plus difficile. La teneur en lignine de la plupart des résidus végétaux pourrait être plus importante que le rapport C:N pour prédire la vitesse de décomposition.
Les champignons mycorhiziens vivent dans le sol, à la surface ou dans les racines des plantes. Les champignons ont une grande surface et contribuent au transport des nutriments minéraux et de l’eau vers les plantes. Le cycle de vie des champignons est plus complexe et plus long que celui des bactéries. Les champignons ne sont pas aussi résistants que les bactéries et nécessitent une source de nourriture plus constante. Les niveaux de population de champignons ont tendance à diminuer avec le travail du sol conventionnel. Les champignons ont un rapport carbone/azote plus élevé (10:1 carbone/azote ou 10 % d’azote) mais sont plus efficaces pour convertir le carbone en matière organique du sol. Avec des résidus organiques riches en C:N, les bactéries et les champignons extraient l’azote du sol (voir le graphique sur l’immobilisation nette).
Les protozoaires et les nématodes consomment d’autres microbes. Les protozoaires peuvent se reproduire en six à huit heures, tandis que les nématodes mettent de trois jours à trois ans, avec une moyenne de 30 jours pour se reproduire. Après avoir consommé les bactéries ou autres microbes (riches en azote), les protozoaires et les nématodes libèrent de l’azote sous forme d’ammonium (voir le graphique sur la minéralisation nette). L’ammonium (NH 4 +) et les nitrates (NO 3 -) sont facilement convertis dans les deux sens dans le sol. Les plantes absorbent les nitrates d’ammonium et du sol pour se nourrir grâce au réseau mycorhizien des champignons.
Les populations de micro-organismes changent rapidement dans le sol à mesure que les produits SOM sont ajoutés, consommés et recyclés. La quantité, le type et la disponibilité de la matière organique détermineront la population microbienne et son évolution. Chaque organisme individuel (bactéries, champignons, protozoaires) possède certaines enzymes et réactions chimiques complexes qui aident cet organisme à assimiler le carbone. À mesure que des déchets sont générés et que les résidus organiques d’origine sont décomposés, de nouveaux micro-organismes peuvent prendre le relais, se nourrissant des déchets, de la nouvelle communauté microbienne florissante (généralement des bactéries) ou de la MOS plus résistante. Les premiers décomposeurs attaquent généralement les sucres et les protéines faciles à digérer, suivis par les micro-organismes qui s’attaquent aux résidus les plus résistants.
Figure 3. Graphique du niébé (C:N <20) décomposé par les bactéries et les champignons, l’évolution du dioxyde de carbone et les protozoaires et les nématodes consommant les bactéries et les champignons et excrétant de l’ammonium dans le sol pour la croissance des plantes. NO3- et NH4+ sont facilement convertis dans le sol. Graphique du Dr Rafiq Islam.
Figure 4. Graphique du niébé (C:N <20) décomposé par les bactéries et les champignons, l’évolution du dioxyde de carbone et les protozoaires et les nématodes consommant les bactéries et les champignons et excrétant de l’ammonium dans le sol pour la croissance des plantes. NO3- et NH4+ sont facilement convertis dans le sol. Graphique du Dr Rafiq Islam.
Les cultures de couverture fournissent de la nourriture (carbone actif comme le glucose et les protéines) aux microbes dont ils peuvent se nourrir. Dans le sol, il y a 1 000 à 2 000 fois plus de microbes associés aux racines que ceux vivant dans un sol nu ou labouré. Les microbes fabriquent à leur tour de la MOS et stockent les nutriments du sol. La construction de SOM nécessite que les éléments nutritifs du sol comme le NPKS soient liés dans le sol. Les cultures de couverture hivernales absorbent l’excès de nutriments du sol et fournissent de la nourriture à tous les microbes présents dans le sol pendant les mois d’hiver, plutôt que d’utiliser les réserves de MOS pour les nutriments. Dans un champ labouré conventionnel, les éléments nutritifs du sol sont rapidement libérés à mesure que la MOS est brûlée et que l’habitat des microbes et des organismes du sol est détruit. Dans un champ sans labour, des niveaux élevés de MOS constituent des réserves de nutriments du sol qui sont lentement libérés dans les sols. L’ajout d’une culture de couverture vivante dans un champ sans labour augmente la matière organique active (sucres et protéines) pour les microbes du sol. Les microbes du sol peuvent se nourrir de deux cultures au lieu d’une seule par an. Les microbes prospèrent dans des conditions de culture sans labour et dans les cultures de couverture hivernales. Les cultures de couverture et le fumier peuvent être utilisés pour nourrir les microbes du sol et recycler les éléments nutritifs du sol. À mesure que les microbes du sol décomposent les résidus organiques, ils libèrent lentement des éléments nutritifs dans le sol pour les cultures de couverture hivernales ou pour la culture précédente. Les cultures de couverture empêchent la perte de nutriments par l’érosion du sol, le lessivage, la volatilisation ou la dénitrification.
Résumé
Les micro-organismes abondent dans le sol et jouent un rôle essentiel dans la décomposition des résidus organiques et le recyclage des éléments nutritifs du sol. Les bactéries sont les microbes les plus petits et les plus résistants du sol et peuvent survivre dans des conditions difficiles comme le travail du sol. Les bactéries ne sont efficaces qu’à 20 à 30 pour cent pour recycler le carbone, ont une teneur élevée en azote (3 à 10 atomes de carbone pour 1 atome d’azote ou 10 à 30 pour cent d’azote), une teneur en carbone plus faible et une durée de vie courte. L’efficacité de l’utilisation du carbone est de 40 à 55 pour cent pour les champignons mycorhiziens, de sorte qu’ils stockent et recyclent plus de carbone (rapport carbone/azote de 10 : 1) et moins d’azote (10 pour cent) dans leurs cellules que les bactéries. Les champignons sont plus spécialisés mais ont besoin d’une source de nourriture constante et se développent mieux dans des conditions sans labour.
La matière organique du sol (MOS) est composée des fractions « vivantes » (micro-organismes), « mortes » (résidus frais) et « très mortes » (humus). La SOM active est composée de matières végétales ou animales fraîches qui servent de nourriture aux microbes et est composée de sucres et de protéines faciles à digérer. La SOM passive est résistante à la décomposition par les microbes (plus riche en lignine). Active SOM améliore la structure du sol et retient les nutriments disponibles pour les plantes. Chaque 1 pour cent de SOM contient 1 000 livres d’azote, 100 livres de phosphore, 100 livres de potassium et 100 livres de soufre ainsi que d’autres nutriments végétaux essentiels. Le travail du sol détruit la MOS en oxydant la MOS, permettant aux bactéries et autres microbes de décomposer rapidement les résidus organiques. Des températures et une humidité plus élevées augmentent la destruction de la MOS en augmentant les populations microbiennes dans le sol. Les résidus organiques avec un faible rapport carbone/azote (C:N) (inférieur à 20) se décomposent facilement et les nutriments sont rapidement libérés (4 à 8 semaines), tandis que les résidus organiques avec un rapport C:N élevé (supérieur à 20) se décomposent. lentement et les microbes vont capter l’azote du sol pour décomposer les résidus. Les protozoaires et les nématodes consomment d’autres microbes présents dans le sol et libèrent de l’azote sous forme d’ammonium, qui devient disponible pour d’autres micro-organismes ou est absorbé par les racines des plantes.
Reconnaissance
Cette fiche d’information a été réalisée en collaboration avec le Midwest Cover Crops Council (MCCC).
Les références
Alexandre, Martin. 1991. Introduction à la microbiologie des sols, 2e éd. Malabar, Floride : Société d’édition Krieger.
L’ ACS expliqué sur l’excellent site triple performance https://wiki.tripleperformance.fr/wiki/Triple_Performance
L’agriculture de conservation des sols (ACS) est un ensemble de techniques culturales destinées à maintenir et améliorer le potentiel agronomique des sols, tout en conservant une production régulière et performante sur les plans technique et économique.
99,99 % Des Pesticides Que Nous Consommons Sont Fabriqués Par Des Plantes
Autant ce que l’on appelle pesticides sont retrouvés NATURELLEMENT jusque dans notre alimentation…..Autant dans la nature vous ne trouverez aucun exemple de travail mécanique de sol comme l’agriculture conventionnelle le pratique . Les plantes n’ont pas besoin de travail mécanique de sol pour se développer …..Mais elles ont besoin de fabriquer des produits pour leurs défenses auprès des prédateurs et des maladies
Quand Bruce Ames parle de toxicité, il est temps d’écouter (1) . Ames est l’inventeur du très important test d’Ames pour la mutagénicité, qui mesure les dommages causés à l’ADN par un produit chimique donné. Le test d’Ames constitue un obstacle essentiel dans le monde de la recherche sur la découverte de médicaments. Même si un test Ames positif ne constitue pas une preuve de facto qu’un produit chimique sera cancérigène chez l’homme, il s’agit d’un signal d’alarme géant dans le développement de médicaments. De nombreux médicaments prometteurs ont rencontré leur fabricant simplement grâce à un test d’Ames positif.
BONNE CHANCE POUR ÉVITER LES PESTICIDES
Étant donné que les pesticides et les herbicides font régulièrement la une des journaux, dernièrement à cause de la « guerre du glyphosate », (2) j’ai pensé qu’il pourrait être intéressant d’examiner un article de synthèse qu’Ames et ses collègues ont écrit il y a près de trois décennies dans les Actes de l’Académie nationale des sciences. (PNAS) . Si vous vivez dans l’illusion que vous pouvez vous lancer dans des aliments entiers et payer trop cher pour un tas de produits biologiques sans pesticides, cet article vous détrompera de cette erreur. Si vous ne voulez pas consommer de pesticides, alors vous feriez mieux d’arrêter de manger, car vous en consommez à chaque bouchée. Il y en a beaucoup. Selon Ames :
99,99 % (en poids) des pesticides consommés par le public américain sont fabriqués par les plantes elles-mêmes comme mécanisme de défense.
Les pesticides synthétiques et naturels sont également susceptibles d’être cancérigènes.
(Il est peu probable que vous voyiez cette information publiée dans Whole Foods.)
Ames écrit (c’est moi qui souligne) :
« L’examen toxicologique des produits chimiques synthétiques tels que les pesticides et les polluants industriels, sans examen similaire des produits chimiques présents dans la nature à des fins de comparaison, a généré un déséquilibre à la fois dans les données et dans la perception des dangers potentiels pour les humains . »
Sans blague. À l’American Council, nous crions cela depuis de nombreuses années, pour ensuite être traités de « complices de l’industrie » chaque fois que nous osons qualifier un produit chimique de sûr. Non seulement nous ne sommes pas des complices (3) , mais nous avons toujours eu raison sur un point clé. La distinction entre produits chimiques « naturels » et « artificiels » n’a aucun sens – ce que beaucoup d’Américains ne connaissent pas. L’origine d’un produit chimique n’a pas d’importance. Votre corps ne peut pas dire ce qui est naturel ou synthétique – seulement les propriétés de la matière chimique.
Il est difficile de blâmer les consommateurs ; l’industrie alimentaire biologique et les charlatans d’Internet ont fait un travail magistral en créant un récit selon lequel nous sommes tous empoisonnés par d’infimes quantités de milliers de produits chimiques fabriqués par l’homme, et que le moyen d’éviter cela est d’acheter des produits dits « naturels ». produits car ils ne contiennent pas de produits chimiques. Mais le récit est complètement faux. Excellente commercialisation. Terrible science. Voyons comment Ames est parvenu à ses conclusions.
MODÈLES DE CANCÉROGÉNICITÉ DU RAT ET DE LA SOURIS – DE NOMBREUSES LIMITES
Toutes les données de cancérogénicité contenues dans l’article du PNAS proviennent de modèles de cancer chez le rat et la souris, qui sont notoirement peu fiables pour prédire les cancers humains, mais ce sont les meilleurs que nous ayons. Ames et ses collègues de l’UC Berkeley ont parcouru la littérature à la recherche d’études sur les aliments contenant des pesticides naturels connus. Les chiffres sont énormes. Par exemple, rien que dans le chou, 49 pesticides et leurs métabolites ont été détectés. Parmi ceux-ci, deux produits chimiques, l’acide chlorogénique et l’isothiocyanate d’allyle, ont provoqué des tumeurs chez le rat, mais pas chez la souris (4) . Ames estime que les Américains consomment entre 5 000 et 10 000 pesticides naturels différents.
LES CANCÉRIGÈNES SONT PARTOUT
La revue a également identifié des études dans lesquelles des pesticides d’origine végétale avaient été testés (à fortes doses) (5) pour voir s’ils provoquaient le cancer chez les rongeurs. Cinquante-deux de ces pesticides, tous couramment présents dans divers aliments, ont été évalués de cette manière ; 27 se sont révélés cancérigènes. Plus de la moitié.
« Les pesticides naturels constituent un sous-ensemble important de produits chimiques naturels. Les plantes produisent des toxines pour se protéger contre les champignons, les insectes et les prédateurs animaux. »
Bruce Ames, et. Al.
Les plantes n’existent pas pour servir les humains ; ils sont ici parce qu’ils ont survécu et se sont reproduits. Pour ce faire, ils ont développé la capacité de synthétiser des produits chimiques pour les protéger des prédateurs. Malgré les protestations constantes contre les pesticides, nous n’aurions rien à manger si les plantes ne produisaient pas celles dont elles ont besoin pour se défendre.
ET NOUS EN MANGERONS BEAUCOUP
Notre obsession idiote d’éviter d’infimes quantités de résidus de pesticides sur une pomme semble encore plus idiote lorsque nous examinons les quantités relatives de pesticides naturels et synthétiques que nous consommons. Lorsque la FDA a analysé les aliments pour détecter la présence de produits chimiques synthétiques importants (probablement présents dans l’environnement), 105 résidus chimiques différents ont été détectés dans les aliments (6) . La somme totale de ces 105 produits chimiques (combinés) a été estimée à environ 0,09 mg par personne et par jour, dont environ la moitié sont cancérigènes. En revanche, nous consommons quotidiennement environ 1,5 g (1 500 mg) de pesticides naturels.
Nous concluons également qu’aux faibles doses de la plupart des expositions humaines, les risques comparatifs des résidus de pesticides synthétiques sont insignifiants .
MAIS NOUS NE SOMMES PAS TOUS MORTS
Nous consommons régulièrement des milliers de pesticides, la plupart fabriqués par les plantes, et une quantité bien moindre de résidus de pesticides appliqués sur les cultures. Et pourtant, nous sommes toujours là. La raison est évidente, et c’est la même raison pour laquelle il n’y a pas de quoi s’inquiéter des infimes quantités de croque-mitaines environnementaux omniprésents comme le BPA, les phtalates et les parabènes : la dose. Les produits chimiques cancérigènes, qu’ils soient d’origine végétale ou synthétiques, sont traités par le foie et excrétés. C’est le travail du foie, et il le fait très bien. Sinon, il n’y aurait pas besoin de pesticides de synthèse car le chou nous aurait déjà planté en terre.
REMARQUES:
(1) Bruce Ames, l’un des fondateurs de l’American Council, n’a pas remporté le prix Nobel pour son invention. Cela me laisse perplexe.
(2) Même si cela n’a rien de drôle, il y a eu beaucoup d’affaires amusantes concernant le glyphosate ces derniers temps, y compris des fraudes. En parlant de fraude, voyez le démantèlement brutal de la corruption du CIRC par mon collègue Alex Berezow . Le Dr Berezow a également écrit ici sur les pesticides à base de plantes .
(3) Des compères ? À peine. À l’ACSH, nous gardons un mur entre la science et la collecte de fonds pour éviter ne serait-ce que l’apparence d’un conflit d’intérêts. À l’heure actuelle, c’est un mur inutile. On m’a dit que 97 % de notre financement provenait de donateurs individuels. Je ne sais pas d’où viennent les 3% restants.
(4) Le statut réglementaire actuel de l’isothiocyanate d’allyle n’est pas cohérent avec les conclusions d’Ames. Le produit chimique n’apparaît pas sur la liste de la proposition 65 de Californie et fait partie du groupe 3 du CIRC – non classifiable quant à sa cancérogénicité pour l’homme.
(4) Cet écart est expliqué par le Programme national de toxicologie car les souris ne reçoivent pas la dose maximale tolérée des deux produits chimiques alors que les rats la reçoivent.
(5) Je ne plaisante pas sur les fortes doses. Environ la moitié de ces tests ont été réalisés à la dose maximale tolérée (DMT) – la dose à laquelle les animaux commencent à mourir si elle est dépassée. On ne sait pas si cela a à voir avec le fait que les humains consomment de petites quantités de ce produit chimique – une autre limite des tests de cancérogénicité sur les rongeurs.
(6) Conseil national de recherches, Conseil de l’agriculture (1987) Réglementation des pesticides dans les aliments (National Academy Press, Washington, DC).





















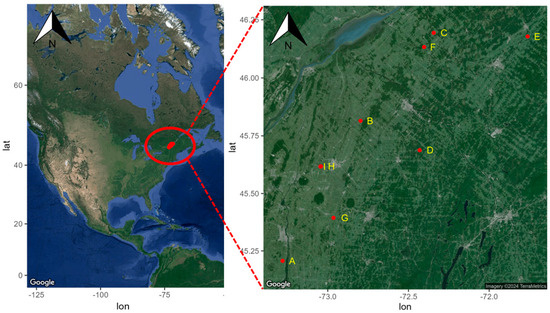
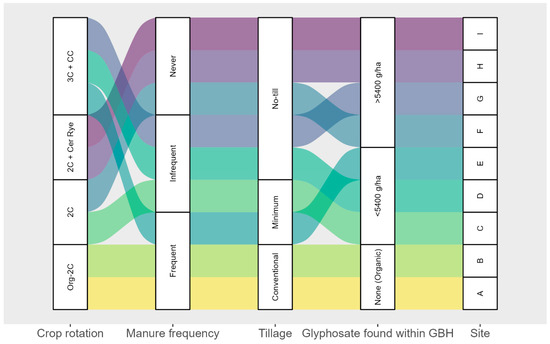
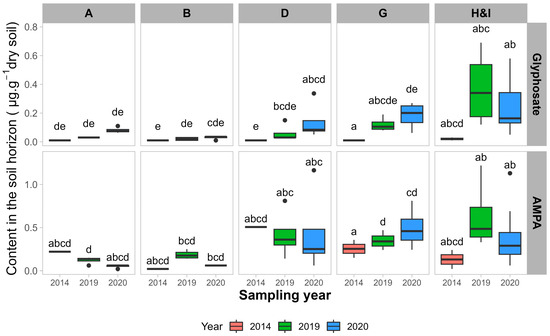
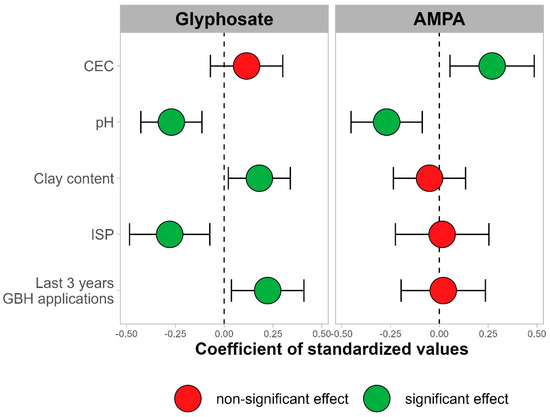
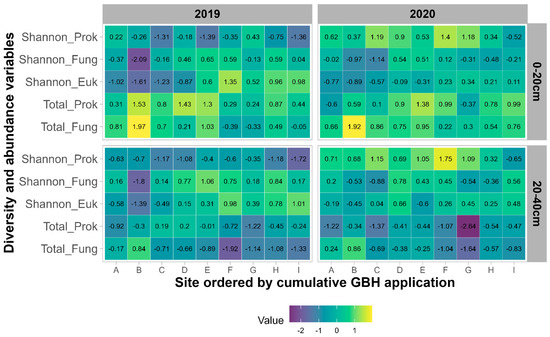
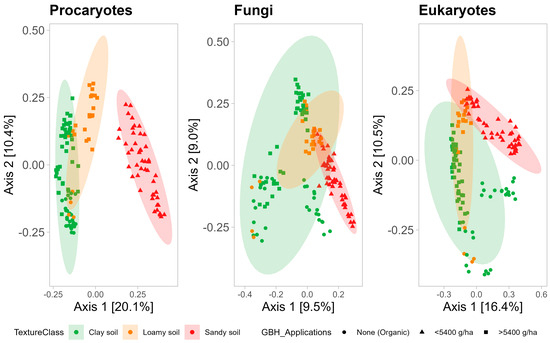
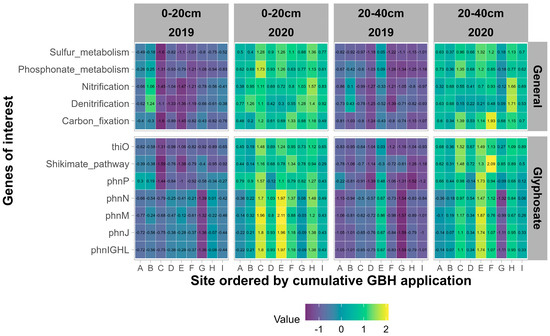


 décomposé par les bactéries et les champignons, l'évolution du dioxyde de carbone et les protozoaires et les nématodes consommant les bactéries et les champignons et excrétant de l'ammonium dans le sol pour la croissance des plantes. NO3- et NH4+ sont facilement convertis dans le sol. Graphique du Dr Rafiq Islam.")
 décomposé par les bactéries et les champignons, l'évolution du dioxyde de carbone et les protozoaires et les nématodes consommant les bactéries et les champignons et excrétant de l'ammonium dans le sol pour la croissance des plantes. NO3- et NH4+ sont facilement convertis dans le sol. Graphique du Dr Rafiq Islam.")
{kind=link}
{kind=link}
{kind=link}
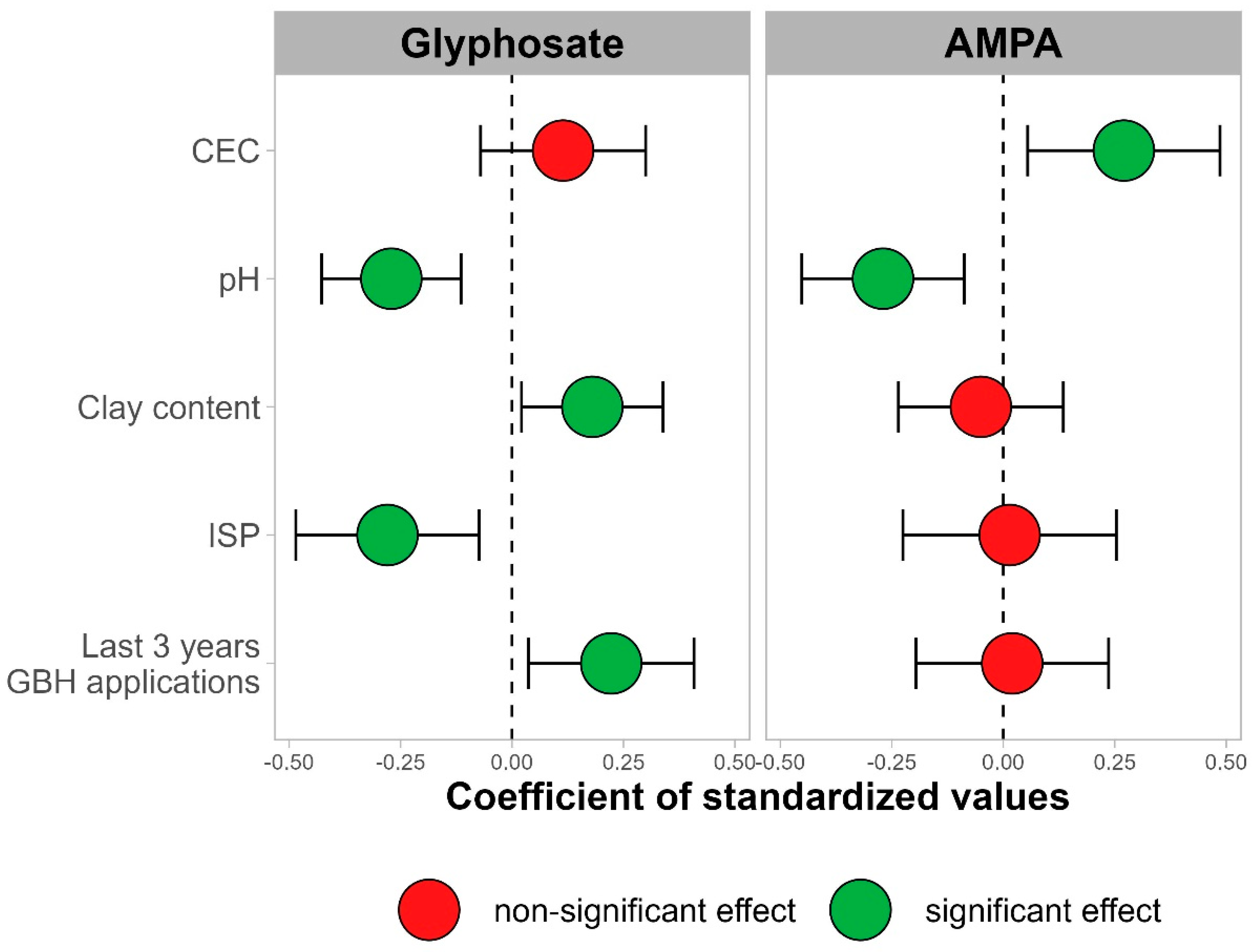{kind=link}
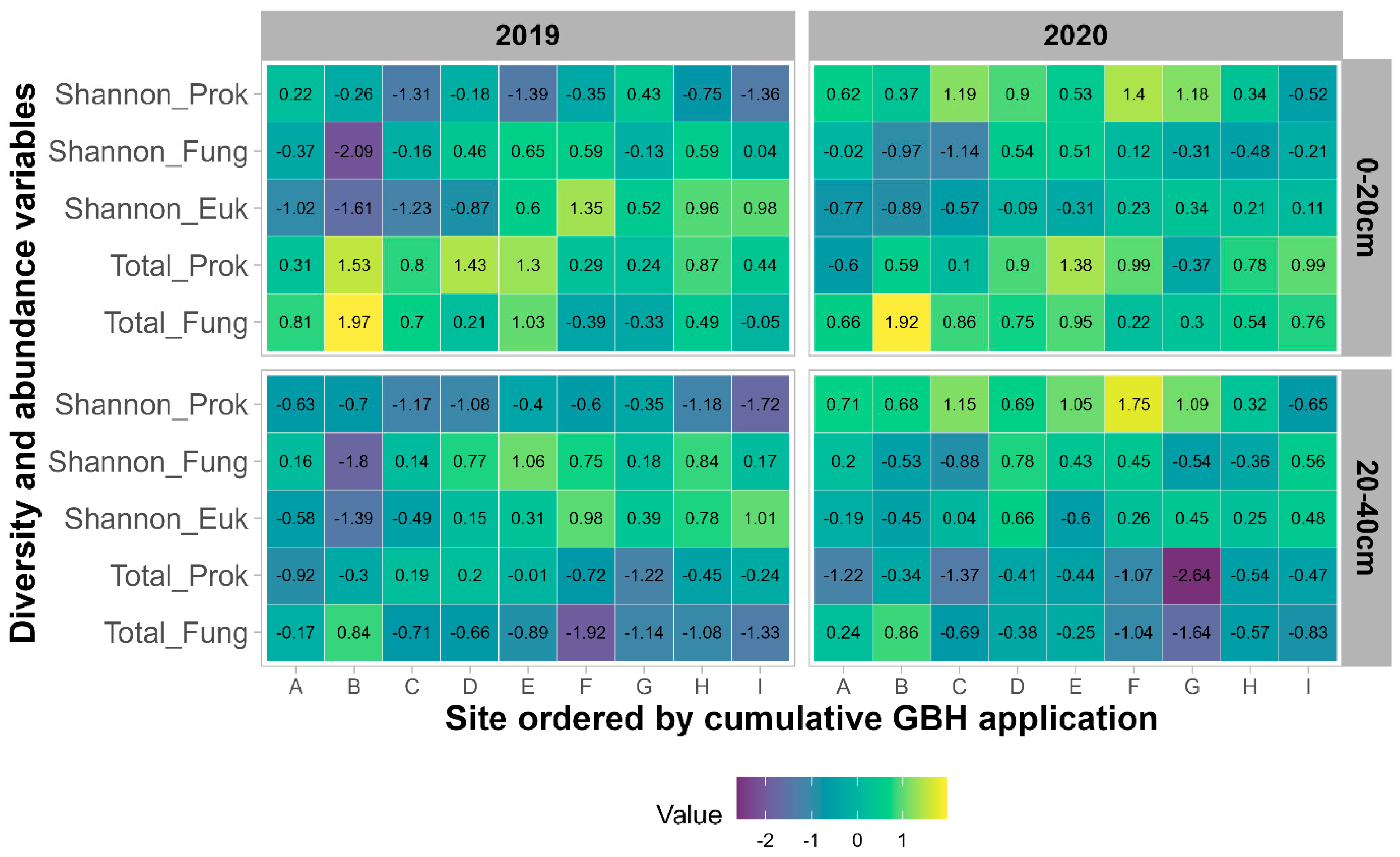{kind=link}
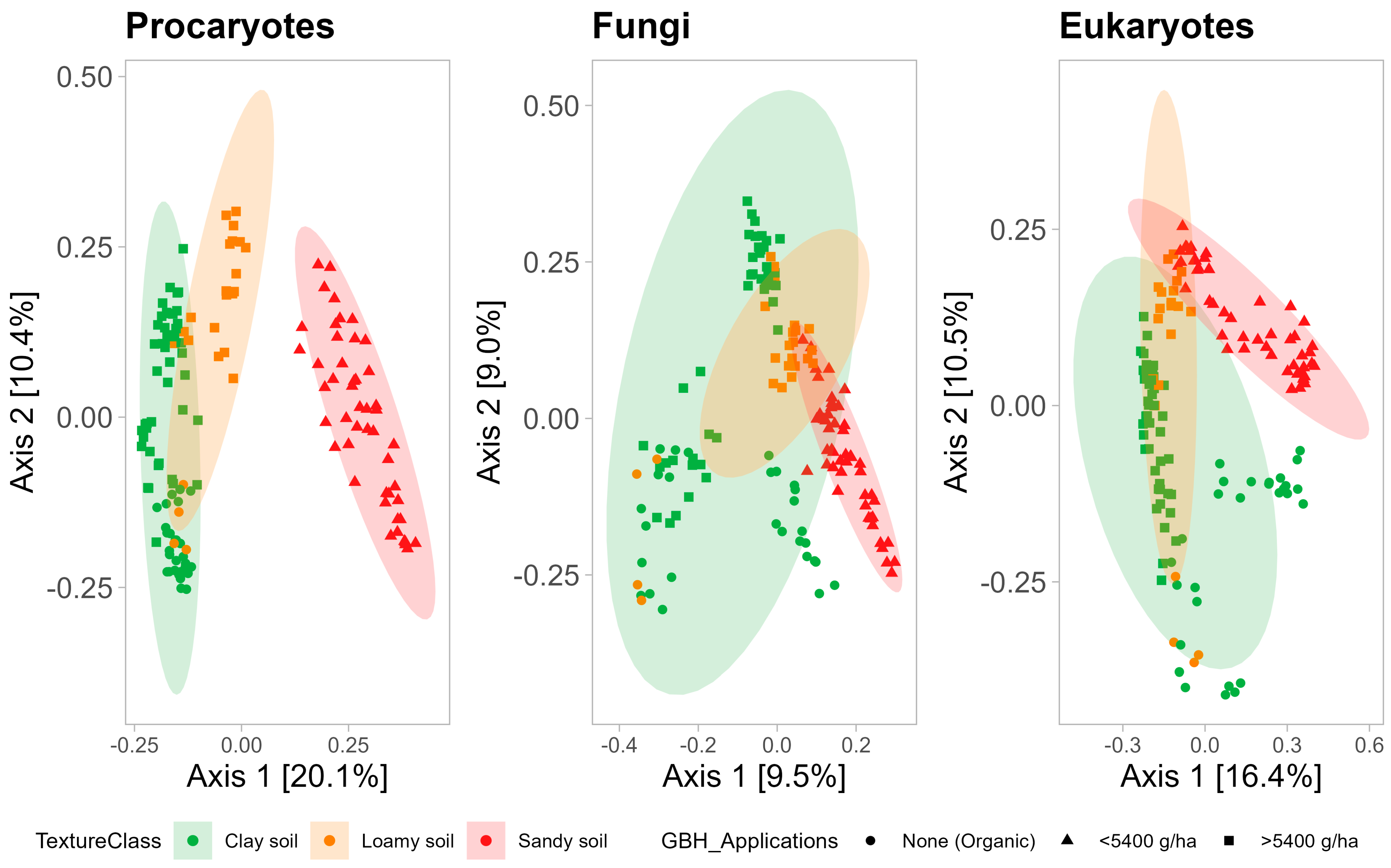{kind=link}
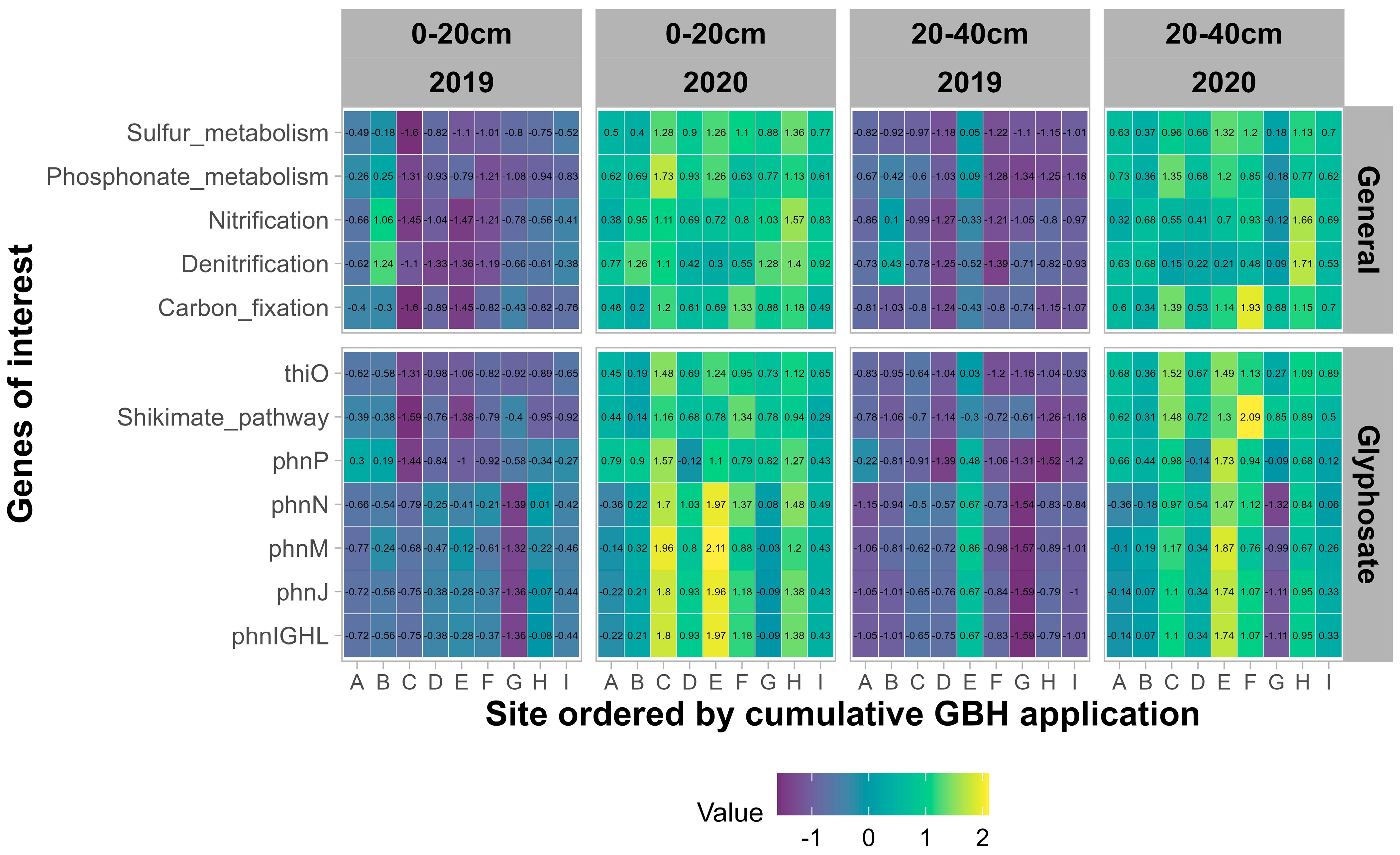{kind=link}