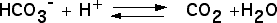Le Centre National d’Agroécologie (CNA) est un projet porté par des acteurs de l’agroécologie qui y fédèrent leurs compétences et expertises : diffusion et échanges de savoirs, formation et accompagnement technique, outils d’évaluation et de mesure pour concevoir et piloter les systèmes agroécologiques. La raison d’être du CNA est de faire émerger des innovations basées sur les principes de l’agroécologie, au service de l’agriculture et des agriculteurs, pour aider à l’évolution et à la diffusion de leurs pratiques.
Produire durablement une nourriture de qualité
Sécuriser le revenu des agriculteurs
Couvrir et nourrir le sol pour protéger l’environnement
Atténuer le changement climatique
Stopper le gâchis d’énergie et ressources
Produire de la biodiversité
Limiter les pollutions systémiques
Créer du sens et de la solidarité
Copier la nature Toujours couvert/jamais travaillé
Recherche et formation …..sont des bases solides pour l’avenir
Produire et redonner du pouvoir aux agriculteurs ? C’est le pari de Lucien Séguy, « agronome », mais surtout « paysan ». Lucien Séguy dédiera sa vie à la fertilité et au « génie végétale ». Sans dogmatisme, il associera la plus grande diversité biologique possible et les techniques agro-industrielles de pointes, créant lui-même plusieurs variétés de semences de riz à haut-rendements.
Les militants de Solidarité & Progrès ont dénoncé une politique de la famine depuis la fin des accords de Bretton Woods (1971) et la publication du rapport « Halte à la croissance » (1972). Depuis lors, la dérégulation met tous les agriculteurs en compétition, accroissant les inégalités ; et l’idéologie décroissante des institutions financières s’oppose à l’équipement de l’agriculture, même là où elle est la seule ressource.
Est-il encore possible (écologiquement ? ) d’augmenter la productivité agricole ? Dans ce dossier, nous vous proposons de suivre le chercheur Lucien Séguy, à la découverte d’un monde longtemps passé inaperçu, celui des sols vivants. Nous verrons comment l’alliance des sols et des agriculteurs peut vaincre la pauvreté et la faim.
Biographie : Une vie au service de la science ; la science au service des paysans
Lucien Séguy en mission d’appui au Maroc. Source : Fert
Né en 1944 dans une famille paysanne de Dordogne, il est le seul de sa fratrie à poursuivre des études supérieures. Diplômé de l’Ecole Nationale Supérieur d’Agronomie de Toulouse, il se spécialise dans la science des sols (pédologie), puis part en service civique au Sénégal en 1967, développer la riziculture paysanne dans la région de la Casamance. Le Centre de coopération internationale en recherche en agronomique pour le développement (Cirad) lui confie sa première affectation professionnelle au Cameroun. Il y met en évidence l’importance de la gestion du sol pour lutter contre la pyriculariose, un champignon qui détruit les feuilles du riz.
1977 : repéré pour ses travaux sur les cultures de riz, il est appelé au Brésil. Débute sa véritable aventure scientifique. Il y travaillera jusqu’à sa retraite en 2009. En parallèle, il lance des missions d’appui aux paysans dans une dizaine de pays, dont Madagascar, la Côte d’Ivoire, le Gabon, le Cameroun, le Sénégal, la Tunisie, le Viêtnam, le Laos, le Cambodge, le Canada, la France métropolitaine et d’outre-mer, jusque dans les mois qui précèdent son décès en avril 2020.
Le texte qui suit est adapté librement du rapport de Lucien Séguy, son ami Serge Bouzinac, et leurs collègues de la recherche brésilienne : « La symphonie inachevée du Semis Direct dans le Brésil central », rédigé en 2008 pour le Cirad et l’Embrapa (la Société Brésilienne de Recherche Agricole).
Les effets spectaculaires de l’érosion en Europe. Source : agriculture-de-conservation.com / C. Henricot
Lorsque Lucien Séguy arrive au Brésil, il rencontre des paysans ruinés par la dégradation du sol. En cause, le labour.
Le labour est pourtant une méthode qui a fait ses preuves en agriculture. Depuis sa découverte dans le croissant fertile, il y a 10 000 ans, plusieurs civilisations agraires y ont eu recours, de la Mésopotamie à l’Europe et l’Asie. Le labour a permis à des centaines de générations de se nourrir, jusqu’à nous.
Il empêche l’installation des mauvaises herbes et des maladies du sol. Mais exige d’énormes quantités de labeur des paysans. Avec un risque de désertification, particulièrement rapide sous les tropiques.
En climat tropical, les sols ont un aspect minéral, comme la terre de remblai peu fertile de nos terrains vagues. Seuls les cinq premiers centimètres de surface sont noirs et fertiles, comme le terreau ou le compost.
Cette mince couche ne peut s’épaissir, car l’humidité et la température tropicale accélèrent tous les processus du vivant : la croissance des plantes cultivées, comme la décomposition de la matière morte au sol. La « matière organique », noire et fertile, est donc fragile sous ces latitudes.
Dans ce contexte, la technique du labour, qui apporte beaucoup d’oxygène au sol, accélère la décomposition de la matière organique, par les microorganismes du sol. C’est « l’érosion invisible ». De plus, mis à nu et cassé en mottes, le sol est emporté dans les fleuves par les puissantes pluies tropicales. C’est « l’érosion visible ».
En forêt, rien ne se perd !
Tout comme Louis Pasteur, Lucien Séguy a passé une partie de sa jeunesse à peindre, exerçant et développant ses talents d’observateur. En grand scientifique, il interroge sans cesse la nature : « Mon maître, mon juge, mon centre d’inspiration principal, c’est la nature, sous sa complexité la plus grande. Vous avez une question à poser ? Posez-la à la nature. Et elle vous répondra ! Il faut simplement faire quelques manip’ pour qu’elle vous réponde. Et le faire de manière scientifique pour comprendre pourquoi cette réponse ; ce qu’elle veut dire exactement. »
Le but de Séguy est d’inventer une agriculture inspirée de la fertilité naturelle des forêts. Il fonde les principes de cette nouvelle agriculture sur quelques observations.
Lucien Séguy note le rôle essentiel de la « matière morte » dans la fertilité des forêts tropicales. Les « éléments fertilisants » qui s’enfonceraient dans le « sol minéral » pouvant malgré tout être recyclés par les racines profondes.
– Une première observation : Le sol forestier n’a pas besoin d’être labouré pour être meuble. Les organismes du sol forestier et les racines des plantes brassent la matière et produisent la porosité nécessaire à la présence raisonnable d’oxygène et d’eau dans le sol forestier.
1er principe : Lucien Séguy propose donc de semer directement les graines dans le sol, sans labour. L’idée n’est pas nouvelle. Ce semis dit « direct » avait permis aux civilisations précolombiennes de prospérer. La méthode en fut oublié, avec leur effondrement. Cette technique est redécouverte aux Etats-Unis dans les années 60, pour parer aux tempêtes de sable ; suite au « Dust Bowl » des années 30, lors de la grande dépression, qui a profondément meurtri le pays, comme l’illustre le film Les raisins de la colère.
C’est le glyphosate, bêtement diabolisé par des écologistes radicaux, qui a permis aux Etats-Unis d’abandonner le labour, donc d’éviter le désert… Le glyphosate agit en bloquant la photosynthèse des mauvaises herbes, rendant caduc l’archaïque labour. À ce titre, Lucien Séguy propose avec humour que soit décerné « le prix Nobel de la paix à Monsanto, pour sa contribution à la préservation des sols ».
Mais – attention – Lucien Séguy est lucide quant au risque de destruction de l’agriculture par les biotechnologies, quand elles sont développées dans un but mercantile. Au sujet de l’herbicide de Monsanto et des OGM, il nous dit : « la productivité n’a pas augmenté depuis leur adoption massive [alors] que les doses d’herbicide sont plutôt en croissance pour contrôler les [plantes] devenues résistantes au glyphosate. » Avoir recours à de plus grands volumes, inutilement… le contraire de l’écologie !
En Semis Direct, le sol n’est pas retourné : à l’avant du tracteur, un « rolofaca » ou « rouleau hacheur » couche le couvert végétal au sol ; à l’arrière, le semoir sème directement les graines sous cette litière. Source : paysan-breton.fr / Toma Swan
– Lucien Séguy fait une deuxième observation : Les sols forestiers sont couverts en permanence d’une litière végétale. Sous cette litière, les premiers centimètres du sol sont le siège de la fertilité de la forêt tropicale. En profondeur, les éléments nutritifs se font beaucoup plus rares !
2nd principe : Pour avoir un sol aéré, riche en nutriments et protégé contre les pluies diluviennes, Lucien Séguy comprend qu’il doit restituer une partie de la culture au sol. L’enjeu est notamment de nourrir les vers de terre et leur cortège d’êtres vivants. Il choisit de recourir à de « puissantes biomasses » pour couvrir le sol et le régénérer en matière organique, qui confère sa fertilité au sol.
Semis direct dans un couvert de sorgho en Europe. La densité et la vigueur du sorgho en font une excellente « plante géante » de couverture, pour contenir les autres plantes et nourrir le sol. Source : agriculture-de-conservation / Cécile Waligora.
– Une ultime observation permet à cet « ingénieur du végétal » d’identifier la complémentarité des plantes dans les systèmes forestiers : toutes ne prospectent pas l’eau aux mêmes profondeurs ; et certaines plantes peuvent arracher des éléments minéraux au sol, pour les mettre à disposition des plantes de la culture suivante.
3ème principe : Lucien Séguy commence donc un travail qui est au cœur de ce qu’il nomme sa « symphonie inachevée » : Il recherche les meilleures successions et associations végétales, permettant de réduire les dépendances aux fertilisants et aux pesticides, et de maximiser les revenus des agriculteurs.
En rupture avec la monoculture, il propose : « L’incorporation de biomasses de couverture encore plus puissantes et plus diversifiées pour que les couverts végétaux des SCV gagnent en multifonctionnalité gratuite. »
Entre deux cultures commerciales, des couverts végétaux sont semés pour réorienter la biologie, la chimie et la structure du sol. Ici, un mélange de tournesol, phacélie et pois. Source : Ver de terre production
Lucien Séguy appelle sa méthode : « Semis sous Couvert Végétal » (SCV).
Avec son binôme Serge Bouzinac et ses partenaires de la recherche et développement brésiliens, Séguy met en place une méthodologie de « recherche-action » sur « système de cultures pérennes » : le chercheur fait ses essais avec, pour et chez les agriculteurs, sur plusieurs années, pour adapter ses travaux aux contraintes de la production.
Pour ces travaux, Lucien Séguy est nommé docteur « Honoris causa » de l’université de Ponta Grossa, une des meilleures universités agricoles du monde, et “Grand Citoyen” par l’Assemblée Législative de l’Etat du Mato Grosso. Mais le paysan-chercheur tient à rendre hommage aux efforts des agriculteurs brésiliens et des acteurs institutionnels :
« Le Brésil a montré au monde sa capacité à développer en moins de 30 ans une agriculture de conservation que le monde entier admire et lui envie, même si elle est largement encore perfectible comme l’ont démontré nos travaux sur les innovations SCV inspirées du fonctionnement de l’écosystème forestier. »
Ajoutons que, bien que fortement mécanisés, les agricultures du Brésil, de l’Argentine et des Etats-Unis restent peu productives, car peu irriguées : dans la région du Mato Grosso entre 2001 et 2007, les rendements par hectare sont en moyenne : de 3,5 tonnes pour le coton, 2,9 tonnes pour le soja et 3,4 tonnes pour le maïs. En France, le blé produit 7 tonnes par hectare et le maïs 9 tonnes. Un ami et disciple de Lucien Séguy, Christian Abadie, produit même 14 tonnes de maïs dans le sud-ouest de la France !
Pourquoi les agriculteurs des Etats-Unis, de l’Argentine et du Brésil acceptent-ils de produire si peu par hectare ? La réponse est en partie économique.
Regardons plutôt, pour le moment, les résultats de la méthode du « Semis sous Couvert Végétal » de Lucien Séguy.
Bilan des « Semis sous Couvert Végétal »
Amélioration du statut social des paysans, accomplissements économiques et environnementaux divers ; les résultats présentés ci-dessous sont non exhaustifs, mais sont l’occasion d’illustrer les problématiques de l’agriculture.
Résultats du groupe agro-industrielMAEDA : Augmentation de la productivité du soja de 25%, et du coton de 45%. Soit une productivité respectable de près de 4 tonnes de soja par hectare et 5 tonnes de coton. Passage à 3 cultures sur 2 ans au lieu de 2. Les marges ont été multipliées par 3. Le nombre de machines agricoles a été réduit de moitié. Le nombre de prestataires de services a chuté de 71%. Et la consommation de carburant a diminué de 70%. Produire plus avec moins !
Des engrais renouvelables : Le manque d’azote et de phosphore dans le sol est ce qui limite le plus la productivité des champs. L’azote constitue 78% de l’air que nous respirons. Il est obtenu industriellement par la réaction du gaz naturel avec l’air. Quant au phosphore, il est aujourd’hui extrait des carrières de « guano », sites de déjections des oiseaux marins, ressource peu renouvelable.
A titre d’exemple, Lucien Séguy introduit la plante sauvage Stylosanthes, pour sa capacité à fixer l’azote atmosphérique et à mobiliser le phosphore du sol. Cet exploit naturel est dû à des symbioses racinaires, avec des bactéries pour l’azote, et avec des champignons pour le phosphore.
En plus de la Stylosanthes, il choisit la Brachiaria, pour ses racines profondes qui récupèrent les nutriments perdus, de même que pour la densité de son couvert au sol et pour sa forte productivité. Lucien Séguy sème ces deux plantes entre les pieds du maïs qui précède la culture de soja. Il en tire une hausse de productivité pour le maïs et le soja, et s’en sert comme fourrage pour l’élevage.
Une réduction des dégâts : Sur soja, cette fertilité organique permet de réduire le nombre de traitements fongicides des 2/3, voire totalement, et d’obtenir des grains de 1 à 3% plus riches en protéines.
Sur coton, en réduisant de 30 à 50% les apports chimiques d’azote et de potassium, en simple complément de la fertilité naturelle du sol, les dégâts liés au ravageur sont nettement plus faibles. Les passages de pesticides peuvent alors être réduits d’1/3.
Une auto-épuration naturelle : Le sol vivant est un « biodigesteur » des résidus végétaux, mais aussi des pesticides, qu’il réduit en molécules de plus en plus simples. Plus il y a de matière végétale au sol, plus les pesticides restent piégés longtemps dans ce biodigesteur, et donc meilleure est la destruction de ces polluants.
En culture de coton, qui exige souvent de nombreux traitements chimiques, les Semis sous Couvert Végétal (SCV) permettent de réduire le volume d’insecticides de près d’un tiers et le nombre de molécules différentes de 18 à 5. Le volume de fongicides (contre les champignons) est réduit de moitié. La fonction auto-épuratrice des sols vivants permet de réduire sous le seuil de détection l’ensemble des 150 molécules et sous-produits polluants recherchés.
Une valorisation du territoire : Vous voulez sauver la forêt amazonienne ? 1/5ème des surfaces déboisées de l’Amazonie sont à l’abandon, car dégradés par le travail du sol. Les SCV peuvent régénérer ces 16,5 millions d’hectares :
« Les SCV les plus puissants permettent, en 3 à 5 ans, de retrouver les teneurs en matière organique et les caractéristiques de distribution des tailles d’agrégats des sols originels sous forêt. »
Autres sols valorisables : les savanes arbustives du Brésil, appelées Cerrados, réputés incultes. Ces terres, qui rappellent nos garrigues méditerranéennes, offrent plus de 50 millions d’hectares faciles à cultiver en SCV.
Avec la régénération des sols forestiers détruits et la valorisation des sols de savane incultes, Lucien Séguy se réjouit que le Brésil puisse doubler ses surfaces de production sans détruire la forêt amazonienne !
Le bocage Normand. Les haies et les arbres créent un microclimat à l’échelle des parcelles. Auteur : Bournagain.
Enfin, l’eau étant un élément essentiel à la vie, Lucien Séguy recommande de maintenir un maillage continu de végétation native de 20 à 50 mètres autour des parcelles, plutôt que des îlots forestiers. Cette « forme géométrique du défrichement » permet d’éviter l’ « effet de poêle surchauffée qui engendre des courants ascendants d’air chaud qui rejettent les pluies vers les forêts galeries à la périphérie ».
Une symphonie “inachevée”
Comme expliqué précédemment, les agricultures de hautes technologies du Brésil, de l’Argentine et des Etats-Unis pourraient assez facilement produire 1 tonne de plus par hectare. Lucien Séguy déplore le manque de formation technique et pointe aussi des raisons économiques.
À l’image des agriculteurs des Etats-Unis, les agriculteurs du Brésil ont adopté le non-labour, poussés par l’érosion des sols et par les prix très bas payés aux productions agricoles. Le non-labour leur permet de réduire leurs dépenses en matériel et essence, et apporte de la durabilité à leurs exploitations.
Au Brésil Central, où 95% des agriculteurs pratiquent le Semis Direct (1er principe : non-labour), le Semis sous Couvert Végétal (2ème & 3ème principes : une végétation puissante et diversifiée) n’est en réalité pratiqué que sur 30% des champs de soja et 15% des champs de maïs.
Ainsi, depuis les années 2000, étant insuffisamment régénérés, les sols du Brésil Central se compactent et un travail superficiel du sol redevient nécessaire. La méthode alors utilisée est le « discage » des 5 premiers centimètres, ce qui constitue une régression vers le labour et la chimie, des surcoûts et une perte de durabilité.
C’est que produire les couverts à aussi un coût ! Les systèmes de SCV nécessitent des investissements de 11% à 29% supérieurs aux simples systèmes sans labour. Hélas, en plus des aléas naturels, le libéralisme rend les prix payés aux producteurs extrêmement fluctuants et généralement plutôt bas. Ainsi, il est probable que les lendemains incertains découragent l’épargne dans le sol.
La formation professionnelle et des marchés organisés apparaissent comme les deux leviers pour qui veut bâtir une politique du sol vivant.
A la frontière des travaux de Séguy, plusieurs disciplines peuvent être convoquées :
Une myriade de naturalistes (dont les spécialisations sont en proportion de la créativité de la Nature) et d’écologues (spécialisés, eux, dans l’étude des interactions naturels) : mobilisés auprès des agronomes (à recruter également !) pour rechercher de nouvelles plantes de couvertures et de nouvelles stratégies d’assemblage du vivant.
Encore une question de moyen ! L’étude des systèmes naturels nécessite des moyens logistiques que l’approche statistique et la modélisation informatique, généralement privilégiées aujourd’hui en écologie scientifique car moins onéreuses, ne devrait qu’appuyer !
Comme pour les plantes que nous mangeons et qui ont été sélectionnées pour être productives, les sélectionneurs peuvent améliorer les caractéristiques des plantes de couverture : Le sorgho blanc sans tanins à haute teneur en protéines, pour farines, bière, papier et amylose ; le sésame qui contient de 50 à 55% d’huile de qualité supérieure, pour les cosmétiques et l’aviation ; le sarrasin, pour les farines sans gluten, …
L’acclimatation de nouvelles plantes de cultures est aussi un moyen d’améliorer la santé du sol. Par exemple, l’intégration du riz pluvial dans les rotations avec les cultures de soja et coton permettrait de lutter contre les maladies à champignon. Pour rendre cette culture de riz pluvial attractive, le défi est actuellement de mettre au point des variétés à hauts rendements.
Quant aux biologistes et aux généticiens, leurs connaissances à l’échelle cellulaire et moléculaire est une aubaine pour l’étude du sol vivant :
Le séquençage génomique, d’abord, pour l’étude de la diversité microbienne. Champignons et bactéries cachent d’innombrables possibilités de symbioses et d’effets antibiotiques naturels. Il s’agit de tirer parti du haut potentiel des sols vivants !
La recherche dans les OGM aussi, à condition d’être au service de l’agronomie, seule discipline globale des champs : « Les SCV offrent une biodiversité fonctionnelle très efficace […] dans un tel contexte, les OGM pourraient être des auxiliaires précieux, légitimes et incontestablement valorisés pour seulement compléter les services écosystémiques majeurs des SCV. »
Nourrir les BRICS
Lucien Séguy, qui dénonce volontiers le conservatisme français, ne s’étend pas sur la politique du Brésil. Tentons, par nous-même, de comprendre cette très jeune puissance agricole :
L’Embrapa, entreprise d’Etat, a acclimaté le blé, le soja, le maïs et le coton, aux sols et aux climats brésiliens. Son existence depuis 1972 inscrit le Brésil dans une démarche scientifique à long terme et ouvre la possibilité de transferts de compétences avec l’Afrique.
Tracés envisagés du chemin de fer “bi-océanique”.
Bien que le Brésil ait su défendre ses intérêts agricoles devant l’Organisation Mondiale du Commerce, le pays tente dès 2003 de remettre en cause les règles arbitraires imposées par les institutions dévoyées de Bretton Woods. En 2014, au sommet de Fortaleza, est lancée la Nouvelle Banque de Développement des BRICS (Brésil, Russie, Inde, Chine et Afrique du Sud).
Il s’agit pour les pays émergents de réduire leurs coûts de production, non par le dumping, mais par l’apport technologique et l’aménagement du territoire. Cette politique est aussi connue sous le nom de « Nouvelles Routes de la Soie ». Un exemple de coopération agricole entre la Chine et le Brésil : la construction de la voie ferrée bi-océanique, traversant l’Amérique latine d’Est en Ouest. Projet gagnant/gagnant, puisqu’il désenclave l’Amazonie et évite le passage par le canal de Panama pour fournir la Chine en denrées agricoles.
En France, les réseaux de Lucien Séguy ont débuté 30 ans après le bio, mais 15% des surfaces agricoles françaises sont déjà cultivées en Semis-direct, soit le double de l’agriculture biologique. En dehors des radars de la plupart des médias.
Bien sûr, le bio nourrit aussi les gens et doit être soutenu à ce titre. Notons seulement ses limites pour mieux nous détacher de l’agribashing : Une productivité moindre et un sol moins riche en vie.
Apprécions aussi les vrais mérites du bio : une production fraîche, car locale, et des circuits-courts, pour que la valeur aille aux producteurs.
Lucien Séguy entouré de Français passionnés. Source : festival Paysage in Marciac, 2018.
Les réseaux français de Lucien Séguy tentent toutes les dénominations pour se faire connaître du public : agriculture de conservation, agriculture de régénération, agriculture sur sol vivant. Et le ver de terre comme symbole de la méthode.
Ces agriculteurs sont présents dans tous les syndicats. Et la Coordination Rurale, second syndicat agricole de France, entre la FNSEA et la Confédération Paysanne, organise depuis 25 ans des festivals de non-labour.
Un ministre, le socialiste Stéphane Le Foll, a rencontré les leaders de ces mouvements en France, mais son cabinet n’a rien trouvé de mieux à leur proposer qu’une tribune à la COP21. Car la priorité du ministère est d’atteindre la neutralité carbone de notre agriculture, d’ici à 2050. Notre article « Ces milliardaires « écolos » qui veulent nous affamer » dénonce cette politique agricole malthusienne.
Une rencontre fortuite nous avait d’ailleurs permis d’interroger le ministre sur la détresse des agriculteurs et les prix trop bas. Nous lui demandions de mettre en place une limitation des importations et un stockage des surplus pour hausser les prix. Réponse du ministre : « Nous ne sommes pas communistes ! » Ce fut l’occasion pour nous d’évoquer la méthode des trente glorieuses : « Monsieur le ministre, De Gaulle non plus ; pourtant, c’était sa politique ».
Stéphane Le Foll a tout de même soutenu la création de « ver de terre production ». Cette chaîne de diffusion de contenus sur Youtube est animée par des agriculteurs. Mais les médias traditionnels restent en roue libre… et le public avec eux. Le pays a-t-il besoin d’une chaîne de télévision publique, qui soit dirigée par les instituts scientifiques ?
Parmi les mesures pour encourager l’agriculture sur sol vivant, rappelons l’importance d’autoriser le glyphosate pour maîtriser les couverts végétaux.
Enfin, l’hexagone gagnerait beaucoup à mettre en place une taxe sur les importations de soja. Elle inciterait le retour de la production de légumineuses en France, comme durant l’année 1973 (lorsque les Américains ont organisé un embargo du soja contre l’Europe).
Qu’est-ce qu’une légumineuse ? Une légumineuse est une plante qui, comme le pois, la lentille ou le soja, est capable de fixer l’azote présent dans l’air. Elles permettent de fertiliser un champ pour la culture suivante. Les légumineuses servent à diversifier les cultures pour limiter l’installation de maladies du sol. Et leurs graines, riches en protéines, augmentent l’autonomie de l’élevage. Cette taxe est donc une mesure de « protectionnisme éducateur », dans la tradition de l’économiste Friedrich List.
Le Brésil a prouvé qu’il est capable d’entendre cet argument, à condition que son droit au développement soit pris en compte. Retrouvons l’état d’esprit des coopérants, comme Lucien Séguy.
Antoine Beils, mai 2021
Annexe I : De l’importance de la productivité « physique » pour nourrir le monde
L’économiste et homme politique américain Lyndon Larouche a montré la relation entre d’une part, la productivité par travailleur, unité de surface et volume de matières utilisées (les 3 paramètres de la productivité physique) contraints par les handicaps naturels d’un territoire ; et d’autre part, le potentiel démographique de ce même territoire. Dit autrement : Le nombre d’humains que la Terre peut porter n’est pas déterminé par les ressources naturelles, mais par l’efficacité de leur utilisation.
Examinons la productivité physique de l’agriculture.
L’agriculture raisonnée, l’agriculture de précision et l’agroécologie permettent toutes trois de produire autant, avec de moindres volumes de produits chimiques. Ce qui n’est pas forcément le cas du bio, qui a globalement recours à de grands volumes de pesticides « naturels ».
Même constat, avec la faible production du bio par hectare et par travailleur ; bien que l’émergence de pratiques agroécologiques (lorsqu’elles sont compatibles avec le cahier des charges du bio) permettent quelques améliorations.
L’Organisation des Nations Unies (ONU) et sa branche pour l’Alimentation et l’Agriculture (FAO) soutiennent cette productivité physique. Elles parlent d’ailleurs d‘ « agriculture écologiquement intensive » ; mais sans moyens financiers à long terme pour promouvoir les semences à hauts rendements, la mécanisation et les autres moyens logistiques du stockage, du transport et de la chaîne du froid. D’ailleurs, le Programme Alimentaire Mondial des Nations Unies cherche encore 5,5 milliards de dollars, pour l’aide alimentaire d’urgence en 2021.
L’écologie, c’est quand les gros mangent les petits ?
Mais si les prix tombent trop, les agriculteurs renoncent à lutter contre les maladies. Au risque de perdre la production…
C’est aussi le risque pris par le bio, qui, en renonçant à la chimie de synthèse, produit globalement un tiers moins de nourriture que l’agriculture conventionnelle à l’hectare, mais est mieux valorisé commercialement.
La page du productivisme, tant décrié encore, a donc été tournée de force ; bien que les extrémistes verts ne s’en satisfassent pas et exigent l’abandon total des équipements motorisés et de la chimie.
Et le pouvoir de nourrir change de main :
1/ Dépossession des agriculteurs. Puisqu’il faut acquérir du foncier, il faut s’endetter au-delà de ce qui est remboursable. Un problème pour la transmission aux jeunes, mais pas un problème pour le rachat par des fonds financiers.
2/ Concentration de l’agro-industrie. À l’image des abattoirs géants, qui imposent leurs conditions commerciales aux éleveurs.
Annexe II : Puit de carbone & pompe à fric
En climat tropical, le Semis Direct sous puissante biomasse permet de produire de 23 à 32 tonnes de matière sèche, aérienne et racinaire, par hectare. Or, cette matière sèche est faite pour moitié de carbone. Si l’intégralité de cette biomasse est restituée au sol, alors 900 kilos à 2 tonnes de carbone sont séquestrés dans ce sol chaque année, et jusqu’à une dizaine d’années de suite.
Comme développé dans le dossier « Le New Deal vert : sortir du piège de la finance verte », la finance verte tente de mettre les États sous tutelle au nom du climat. Pour l’agriculture, aussi, accepter l’agenda “zéro carbone” est dangereux, car cet agenda mène à l’abandon des énergies fossiles, de la fertilisation azotée et de l’élevage, pourtant nécessaires pour nourrir correctement 9 milliards d’êtres humains en 2050.
Annexe III : Une plongée dans le sol vivant
Voir la vidéo : « Lucien Séguy, semis direct sur 20 millions d’hectares ». École Nationale Supérieure Agronomique de Toulouse (ENSAT).
Arnaud Beils says:Merci pour cet excellent article où j’ai appris beaucoup de choses sur l’intérêt du Semis direct sur Couvert Végétal. Je note notamment la capacité de certaines plantes à aller chercher les minéraux en profondeur. A noter également l’importance méconnus de cette pratique en France (15% de notre production !) Peut-être le semis sous couvert végétal permet-il aussi de réaliser des économies d’eau grâce au fait que le couvert végétal conserve l’humidité contrairement à de la terre nue ?Je souhaiterais te poser quelques questions pour comprendre ce que tu as écrit à fond. 1/ Pourquoi le Brésil n’arrive t’il pas au même rendement que la France ? Tu précises qu’ils sont fortement mécanisés et qu’en 2007, nous (la France) arrivons au double du rendement par hectare. Hypothèse, le sol n’est pas encore assez généré ?2/ L’équivalent du semis direct sous couvert végétal appliqué à de faibles surfaces (- de 1 hectare) est il le maraichage sur sol vivant ? Autrement, quels outils utiliser pour produire en SCV sur une faible surface ?3/ peut-on dire que l’augmentation du CO2 dans l’atmosphère (infime précisons le tout de même) est en parti dût au fait que l’on ne restitue pas la matière organique à la terre ? (d’où un argument supplémentaire en faveur du SCV pour capter le CO2)Encore merci pour cet apport intellectuel. Militons pour que la recherche agronomique ait les moyens d’augmenter notre potentiel démographique. Vive l’Humain explorateur du vivant !Reply
antoinebeils says:Merci à toi l’ami,Pour répondre rapidement : 1/ Comment doubler ou tripler la production : – Le Brésil est un pays aux climats contrastés. Le Mato Grosso par exemple, où a travaillé Séguy, est sec une partie de l’année. Il faut donc investir dans l’irrigation. – Une vraie régénération des sols permet les symbioses racinaires avec la vie du sol, pour valoriser la fertilité, donc une meilleur expression des potentialités de la plante. – Précisions : le soja étant 2 à 3 fois plus riche en protéines que le maïs, les faibles rendements sont compensés.2/ Comment appliquer l’ “agriculture sur sol vivant” sur une surface d’1 hectare : – La chaîne youtube “Ver de Terre Production”, animée par des agriculteurs et des chercheurs, permet de se mettre à jour de “l’état de l’art”, du jardin nourricier aux grandes cultures, en passant par l’élevage, l’arboriculture et le maraichage. 😉3/ La fixation du carbone, un argument pour les sols vivants ? Oui, mais… Le paradigme politique “neutralité carbone en 2050” est dangereux, car il implique la suppression de la fertilisation azoté et le démantèlement de l’élevage, comme l’exigent certains financiers fanatiques. Parlons plutôt de “l’agriculture sur sol vivant” comme d’une méthode pour nourrir le monde, bien nourrir, tout le monde !
James J. Hoorman, cultures de couverture et qualité de l’eau, éducateur en vulgarisation, vulgarisation de l’Ohio State UniversityRafiq Islam, spécialiste des sols et de l’eau, Ohio State University Extension, South Centers at Piketon
Les micro-organismes du sol existent en grand nombre dans le sol tant qu’il existe une source de carbone pour l’énergie. Il existe un grand nombre de bactéries dans le sol, mais en raison de leur petite taille, leur biomasse est plus faible. Les actinomycètes sont 10 fois plus petits en nombre, mais sont plus grands en taille et ont donc une biomasse similaire à celle des bactéries. Les populations de champignons sont plus petites mais dominent la biomasse du sol lorsque le sol n’est pas perturbé. Les bactéries, les actinomycètes et les protozoaires sont robustes et peuvent tolérer davantage de perturbations du sol que les populations fongiques. Ils dominent donc dans les sols labourés, tandis que les populations fongiques et nématodes ont tendance à dominer dans les sols labourés ou sans labour.
Il y a plus de microbes dans une cuillère à café de terre qu’il n’y a d’habitants sur terre. Les sols contiennent environ 8 à 15 tonnes de bactéries, champignons, protozoaires, nématodes, vers de terre et arthropodes. Voir les fiches d’information sur les rôles des bactéries, des champignons, des protozoaires et des nématodes du sol.
Tableau 1 : Nombre relatif et biomasse des espèces microbiennes à une profondeur de sol de 0 à 6 pouces (0 à 15 cm)
Microorganismes
Nombre/g de sol
Biomasse (g/m 2 )
Bactéries
10 8 –10 9
40 à 500
Actinomycètes
10 7 –10 8
40 à 500
Champignons
10 5 –10 6
100-1500
Algues
10 4 –10 5
1 à 50
Protozoaires
10 3 –10 4
Varie
Nématodes
10 2 –10 3
Varie
Décomposition microbienne de la matière organique du sol
La décomposition de la matière organique remplit deux fonctions pour les micro-organismes : fournir de l’énergie pour la croissance et fournir du carbone pour la formation de nouvelles cellules. La matière organique du sol (MOS) est composée des fractions « vivantes » (micro-organismes), « mortes » (résidus frais) et « très mortes » (humus). Le « très mort » ou humus est la fraction de MOS à long terme, vieille de plusieurs milliers d’années et résistante à la décomposition. La matière organique du sol contient deux composants appelés MOS active (35 %) et passive (65 %). La MOS active est composée de matières végétales ou animales fraîches « vivantes » et « mortes » qui servent de nourriture aux microbes et sont composées de sucres et de protéines faciles à digérer. La SOM passive est résistante à la décomposition par les microbes et est plus riche en lignine.
Les microbes ont besoin d’un apport régulier de MOS active dans le sol pour survivre dans le sol. Les sols non labourés à long terme contiennent des niveaux de microbes nettement plus élevés, plus de carbone actif, plus de MOS et plus de carbone stocké que les sols labourés conventionnels. La majorité des microbes présents dans le sol existent dans des conditions de famine et ont donc tendance à être dans un état dormant, en particulier dans les sols labourés.
Les résidus végétaux morts et les éléments nutritifs des plantes deviennent la nourriture des microbes présents dans le sol. La matière organique du sol (MOS) correspond essentiellement à toutes les substances organiques (tout ce qui contient du carbone) présentes dans le sol, vivantes et mortes. La MOS comprend les plantes, les algues bleu-vert, les micro-organismes (bactéries, champignons, protozoaires, nématodes, coléoptères, collemboles, etc.) et la matière organique fraîche et en décomposition provenant des plantes, des animaux et des micro-organismes.
La matière organique du sol peut être décomposée en ses éléments constitutifs. Cent grammes (g) ou 100 livres (lbs) de matière végétale morte produisent environ 60 à 80 g (lbs) de dioxyde de carbone, qui est rejeté dans l’atmosphère. Les 20 à 40 g (lb) d’énergie et de nutriments restants sont décomposés et transformés en environ 3 à 8 g (lb) de micro-organismes (les vivants), 3 à 8 g (lb) de composés non humiques (les morts), et 10 à 30 g (lb) d’humus (la matière très morte, résistante à la décomposition). La structure moléculaire de la MOS est principalement constituée de carbone et d’oxygène avec un peu d’hydrogène et d’azote et de petites quantités de phosphore et de soufre. La matière organique du sol est un sous-produit des cycles du carbone et de l’azote.
Figure 1. Diagramme du Dr Rafiq Islam.
Nutriments de la matière organique du sol
Les éléments nutritifs du sol ont une valeur actuelle de 680 $ pour chaque 1 pour cent de MOS ou de 68 $ par tonne de MOS sur la base des valeurs économiques des engrais commerciaux (voir tableau 2). La MOS est composée principalement de carbone, mais associée au carbone sont de grandes quantités d’azote et de soufre provenant des protéines, du phosphore et du potassium. Le SOM doit être considéré comme un investissement dans un certificat de dépôt (CD). Les sols biologiquement actifs et contenant de plus grandes quantités de carbone actif recyclent et libèrent plus de nutriments pour la croissance des plantes que les sols biologiquement inactifs et contiennent moins de matière organique active. Dans des conditions de culture sans labour, de petites quantités de nutriments sont libérées chaque année (comme les intérêts sur un CD) pour fournir des nutriments lentement et efficacement aux racines des plantes. Cependant, avec le travail du sol, de grandes quantités de nutriments peuvent être libérées puisque la MOS est consommée et détruite par les microbes. Étant donné que les niveaux de MOS mettent du temps à s’accumuler, la capacité de stockage des nutriments est diminuée et les excédents de nutriments libérés sont souvent lessivés dans les eaux de surface. La SOM est un réservoir de nombreux nutriments végétaux.
Considérez les trois scénarios suivants. Les sols renouvellent généralement 1 à 3 pour cent de leur azote stocké dans la MOS. Les sols labourés ou malsains libèrent un pourcentage d’azote plus faible en raison d’une activité microbienne plus faible. Un sol labouré avec 2 pour cent de MOS (2 000 livres de N) peut libérer 1 pour cent de N ou 20 livres de N par an. Un sol plus biologiquement actif et contenant 4 pour cent de MOS (4 000 lb de N) peut libérer 1,5 pour cent de N ou 60 lb de N, tandis qu’un sol à 6 pour cent de MOS (6 000 lb de N) peut libérer 2 pour cent de N ou 120 lb de N. Dans les sols, les excédents de nutriments libérés sont souvent perdus et les réserves de carbone sont épuisées, ce qui réduit le stockage futur des nutriments. Les agriculteurs constatent souvent cela lorsqu’ils labourent un sol vierge, un vieux pâturage ou une clôture. Pendant plusieurs années, les cultures sur le sol nouvellement labouré pousseront mieux que les sols environnants, mais avec le temps, le sol sera épuisé en carbone et le sol nouvellement labouré deviendra moins fertile car le carbone est oxydé sous forme de dioxyde de carbone et perdu dans l’atmosphère. . Le travail du sol entraîne l’oxydation et la destruction du carbone dans le sol en augmentant les niveaux d’oxygène du sol, favorisant ainsi l’expansion des populations de bactéries et la consommation de carbone actif dans le sol.
Tableau 2 : Valeur de la matière organique du sol
Hypothèses : 2 000 000 livres de sol dans les 6 premiers pouces
Nutriments
1% de matière organique = 20 000# 50%Carbone, rapport C:N = 10:1
Effets du climat, de la température et du pH sur la MOS
La MOS est affectée par le climat et la température. Les populations microbiennes doublent à chaque changement de température de 10 degrés Fahrenheit. Si nous comparons les tropiques aux régions arctiques plus froides, nous constatons que la majeure partie du carbone est retenue dans les arbres et la végétation aérienne. Sous les tropiques, la couche arable contient très peu de MOS car les températures élevées et l’humidité décomposent rapidement la MOS. En se déplaçant vers le nord ou le sud de l’équateur, la MOS augmente dans le sol. La toundra près du cercle polaire arctique contient une grande quantité de MOS en raison des températures froides. Les températures glaciales modifient le sol de sorte qu’une plus grande quantité de MOS est décomposée que dans les sols non sujets au gel.
L’humidité, le pH, la profondeur du sol et la taille des particules affectent la décomposition de la MOS. Les régions chaudes et humides stockent moins de carbone organique dans le sol que les régions sèches et froides en raison d’une décomposition microbienne accrue. Le taux de décomposition de la MOS augmente lorsque le sol est exposé à des cycles de séchage et d’humidification par rapport aux sols continuellement humides ou secs. Toutes choses étant égales par ailleurs, les sols dont le pH est neutre à légèrement alcalin décomposent la MOS plus rapidement que les sols acides ; par conséquent, le chaulage du sol améliore la décomposition de la MOS et l’évolution du dioxyde de carbone. La décomposition est également plus importante près de la surface du sol, là où se trouvent la plus forte concentration de résidus végétaux. À de plus grandes profondeurs, la décomposition de la MOS est moindre, ce qui correspond à une baisse des niveaux de carbone organique due à la diminution des résidus végétaux. Les petites particules sont plus facilement dégradées par les microbes du sol que les grosses particules, car la surface globale est plus grande avec les petites particules, de sorte que les microbes peuvent attaquer les résidus.
Une différence dans la formation des sols se produit également d’est en ouest à travers les États-Unis. À l’est, les forêts de feuillus prédominaient et les racines pivotantes des arbres étaient riches en lignine, et les arbres à feuilles caduques laissaient de grandes quantités de feuilles mortes à la surface du sol. Les racines des feuillus ne se renouvellent pas rapidement, de sorte que les niveaux de matière organique dans le sous-sol sont assez faibles. Dans les sols forestiers, la majeure partie de la MOS est distribuée dans les quelques centimètres supérieurs. À mesure que vous vous déplacez vers l’ouest, de hautes prairies dominent le paysage et la couche arable se forme à partir de systèmes racinaires profonds et fibreux. Cinquante pour cent des racines d’une herbe meurent et sont remplacées chaque année et les racines de l’herbe sont riches en sucres et en protéines (matière organique active plus élevée) et plus faibles en lignine. Ainsi, les sols formés sous les prairies à herbes hautes sont riches en MOS dans tout le profil pédologique. Ces sols de premier choix sont très productifs car ils ont un pourcentage plus élevé de MOS (en particulier de charbon actif), retiennent plus de nutriments, contiennent plus de microbes et ont une meilleure structure du sol en raison de populations fongiques plus importantes.
Rapport carbone/azote
La dégradation des résidus organiques par les microbes dépend du rapport carbone/azote (C:N). Les microbes présents dans le rumen d’une vache, dans un tas de compost et dans le sol dépendent du rapport C:N pour décomposer les résidus organiques (à base de carbone). Considérez deux sources d’alimentation distinctes, un jeune plant de luzerne tendre et de la paille d’avoine ou de blé. Une jeune plante de luzerne contient plus de protéines brutes, d’acides aminés et de sucres dans la tige, elle est donc facilement digérée par les microbes, que ce soit dans le rumen d’une vache, dans un tas de compost ou dans le sol. La jeune luzerne a une teneur élevée en azote provenant des protéines (les acides aminés et les protéines sont riches en azote et en soufre), elle a donc un rapport carbone/azote plus faible (moins de carbone, plus d’azote). Cependant, la paille d’avoine et de blé (ou le foin plus âgé) contient plus de lignine (qui résiste à la décomposition microbienne), moins de protéines brutes, moins de sucres dans la tige et un rapport C:N plus élevé. La paille est décomposée par les microbes, mais il faut plus de temps et d’azote pour décomposer cette source riche en carbone.
Une faible teneur en azote ou un rapport C:N élevé est associé à une lente décomposition de la MOS. Les plantes immatures ou jeunes ont une teneur en azote plus élevée, des rapports C:N plus faibles et une décomposition plus rapide de la SOM. Pour un bon compostage, un rapport C:N inférieur à 20 permet aux matières organiques de se décomposer rapidement (4 à 8 semaines) tandis qu’un rapport C:N supérieur à 20 nécessite un supplément d’azote et ralentit la décomposition. Ainsi, si nous ajoutons au sol un matériau à haute teneur en carbone et à faible teneur en N, les microbes capteront l’azote du sol. Finalement, l’azote du sol est libéré mais à court terme, l’azote est retenu. Le facteur de conversion pour convertir l’azote en protéines brutes est de 16,7, ce qui explique pourquoi il est si important d’avoir un rapport C:N inférieur à 20.
Le rapport C:N de la plupart des sols est d’environ 10:1, ce qui indique que l’azote est disponible pour la plante. Le rapport C:N de la plupart des résidus végétaux a tendance à diminuer avec le temps, à mesure que la MOS se désintègre. Cela résulte de la perte gazeuse de dioxyde de carbone. Par conséquent, le pourcentage d’azote dans la MOS résiduelle augmente à mesure que la décomposition progresse. Le rapport C:N de 10:1 de la plupart des sols reflète une valeur d’équilibre associée à la plupart des microbes du sol (bactéries 3:1 à 10:1, champignon 10:1 rapport C:N).
Les bactéries sont les premiers microbes à digérer les nouveaux résidus organiques végétaux et animaux présents dans le sol. Les bactéries peuvent généralement se reproduire en 30 minutes et ont une teneur élevée en N dans leurs cellules (3 à 10 atomes de carbone pour 1 atome d’azote ou 10 à 30 pour cent d’azote). Dans de bonnes conditions de chaleur, d’humidité et de nourriture, ils peuvent se reproduire très rapidement. Les bactéries sont généralement moins efficaces pour convertir le carbone organique en nouvelles cellules. Les bactéries aérobies assimilent environ 5 à 10 pour cent du carbone tandis que les bactéries anaérobies n’en assimilent que 2 à 5 pour cent, laissant derrière elles de nombreux déchets composés de carbone et utilisant de manière inefficace l’énergie stockée dans la MOS.
Figure 2. Graphique du N relatif disponible avec la durée de décomposition.
Luzerne, faible rapport C:N, C:N = 13:1
Paille d’avoine, rapport C:N élevé, C:N = 80:1
Les champignons libèrent généralement moins de dioxyde de carbone dans l’atmosphère et sont plus efficaces pour convertir le carbone pour former de nouvelles cellules. Le champignon capte généralement plus d’énergie de la MOS à mesure qu’il la décompose, assimilant 40 à 55 pour cent du carbone. La plupart des champignons consomment de la matière organique plus riche en cellulose et en lignine, dont la décomposition est plus lente et plus difficile. La teneur en lignine de la plupart des résidus végétaux pourrait être plus importante que le rapport C:N pour prédire la vitesse de décomposition.
Les champignons mycorhiziens vivent dans le sol, à la surface ou dans les racines des plantes. Les champignons ont une grande surface et contribuent au transport des nutriments minéraux et de l’eau vers les plantes. Le cycle de vie des champignons est plus complexe et plus long que celui des bactéries. Les champignons ne sont pas aussi résistants que les bactéries et nécessitent une source de nourriture plus constante. Les niveaux de population de champignons ont tendance à diminuer avec le travail du sol conventionnel. Les champignons ont un rapport carbone/azote plus élevé (10:1 carbone/azote ou 10 % d’azote) mais sont plus efficaces pour convertir le carbone en matière organique du sol. Avec des résidus organiques riches en C:N, les bactéries et les champignons extraient l’azote du sol (voir le graphique sur l’immobilisation nette).
Les protozoaires et les nématodes consomment d’autres microbes. Les protozoaires peuvent se reproduire en six à huit heures, tandis que les nématodes mettent de trois jours à trois ans, avec une moyenne de 30 jours pour se reproduire. Après avoir consommé les bactéries ou autres microbes (riches en azote), les protozoaires et les nématodes libèrent de l’azote sous forme d’ammonium (voir le graphique sur la minéralisation nette). L’ammonium (NH 4 +) et les nitrates (NO 3 -) sont facilement convertis dans les deux sens dans le sol. Les plantes absorbent les nitrates d’ammonium et du sol pour se nourrir grâce au réseau mycorhizien des champignons.
Les populations de micro-organismes changent rapidement dans le sol à mesure que les produits SOM sont ajoutés, consommés et recyclés. La quantité, le type et la disponibilité de la matière organique détermineront la population microbienne et son évolution. Chaque organisme individuel (bactéries, champignons, protozoaires) possède certaines enzymes et réactions chimiques complexes qui aident cet organisme à assimiler le carbone. À mesure que des déchets sont générés et que les résidus organiques d’origine sont décomposés, de nouveaux micro-organismes peuvent prendre le relais, se nourrissant des déchets, de la nouvelle communauté microbienne florissante (généralement des bactéries) ou de la MOS plus résistante. Les premiers décomposeurs attaquent généralement les sucres et les protéines faciles à digérer, suivis par les micro-organismes qui s’attaquent aux résidus les plus résistants.
Figure 3. Graphique du niébé (C:N <20) décomposé par les bactéries et les champignons, l’évolution du dioxyde de carbone et les protozoaires et les nématodes consommant les bactéries et les champignons et excrétant de l’ammonium dans le sol pour la croissance des plantes. NO3- et NH4+ sont facilement convertis dans le sol. Graphique du Dr Rafiq Islam.
Figure 4. Graphique du niébé (C:N <20) décomposé par les bactéries et les champignons, l’évolution du dioxyde de carbone et les protozoaires et les nématodes consommant les bactéries et les champignons et excrétant de l’ammonium dans le sol pour la croissance des plantes. NO3- et NH4+ sont facilement convertis dans le sol. Graphique du Dr Rafiq Islam.
Les cultures de couverture fournissent de la nourriture (carbone actif comme le glucose et les protéines) aux microbes dont ils peuvent se nourrir. Dans le sol, il y a 1 000 à 2 000 fois plus de microbes associés aux racines que ceux vivant dans un sol nu ou labouré. Les microbes fabriquent à leur tour de la MOS et stockent les nutriments du sol. La construction de SOM nécessite que les éléments nutritifs du sol comme le NPKS soient liés dans le sol. Les cultures de couverture hivernales absorbent l’excès de nutriments du sol et fournissent de la nourriture à tous les microbes présents dans le sol pendant les mois d’hiver, plutôt que d’utiliser les réserves de MOS pour les nutriments. Dans un champ labouré conventionnel, les éléments nutritifs du sol sont rapidement libérés à mesure que la MOS est brûlée et que l’habitat des microbes et des organismes du sol est détruit. Dans un champ sans labour, des niveaux élevés de MOS constituent des réserves de nutriments du sol qui sont lentement libérés dans les sols. L’ajout d’une culture de couverture vivante dans un champ sans labour augmente la matière organique active (sucres et protéines) pour les microbes du sol. Les microbes du sol peuvent se nourrir de deux cultures au lieu d’une seule par an. Les microbes prospèrent dans des conditions de culture sans labour et dans les cultures de couverture hivernales. Les cultures de couverture et le fumier peuvent être utilisés pour nourrir les microbes du sol et recycler les éléments nutritifs du sol. À mesure que les microbes du sol décomposent les résidus organiques, ils libèrent lentement des éléments nutritifs dans le sol pour les cultures de couverture hivernales ou pour la culture précédente. Les cultures de couverture empêchent la perte de nutriments par l’érosion du sol, le lessivage, la volatilisation ou la dénitrification.
Résumé
Les micro-organismes abondent dans le sol et jouent un rôle essentiel dans la décomposition des résidus organiques et le recyclage des éléments nutritifs du sol. Les bactéries sont les microbes les plus petits et les plus résistants du sol et peuvent survivre dans des conditions difficiles comme le travail du sol. Les bactéries ne sont efficaces qu’à 20 à 30 pour cent pour recycler le carbone, ont une teneur élevée en azote (3 à 10 atomes de carbone pour 1 atome d’azote ou 10 à 30 pour cent d’azote), une teneur en carbone plus faible et une durée de vie courte. L’efficacité de l’utilisation du carbone est de 40 à 55 pour cent pour les champignons mycorhiziens, de sorte qu’ils stockent et recyclent plus de carbone (rapport carbone/azote de 10 : 1) et moins d’azote (10 pour cent) dans leurs cellules que les bactéries. Les champignons sont plus spécialisés mais ont besoin d’une source de nourriture constante et se développent mieux dans des conditions sans labour.
La matière organique du sol (MOS) est composée des fractions « vivantes » (micro-organismes), « mortes » (résidus frais) et « très mortes » (humus). La SOM active est composée de matières végétales ou animales fraîches qui servent de nourriture aux microbes et est composée de sucres et de protéines faciles à digérer. La SOM passive est résistante à la décomposition par les microbes (plus riche en lignine). Active SOM améliore la structure du sol et retient les nutriments disponibles pour les plantes. Chaque 1 pour cent de SOM contient 1 000 livres d’azote, 100 livres de phosphore, 100 livres de potassium et 100 livres de soufre ainsi que d’autres nutriments végétaux essentiels. Le travail du sol détruit la MOS en oxydant la MOS, permettant aux bactéries et autres microbes de décomposer rapidement les résidus organiques. Des températures et une humidité plus élevées augmentent la destruction de la MOS en augmentant les populations microbiennes dans le sol. Les résidus organiques avec un faible rapport carbone/azote (C:N) (inférieur à 20) se décomposent facilement et les nutriments sont rapidement libérés (4 à 8 semaines), tandis que les résidus organiques avec un rapport C:N élevé (supérieur à 20) se décomposent. lentement et les microbes vont capter l’azote du sol pour décomposer les résidus. Les protozoaires et les nématodes consomment d’autres microbes présents dans le sol et libèrent de l’azote sous forme d’ammonium, qui devient disponible pour d’autres micro-organismes ou est absorbé par les racines des plantes.
Reconnaissance
Cette fiche d’information a été réalisée en collaboration avec le Midwest Cover Crops Council (MCCC).
Les références
Alexandre, Martin. 1991. Introduction à la microbiologie des sols, 2e éd. Malabar, Floride : Société d’édition Krieger.
Vidéo détaillant le pâturage des ovins sur les couverts végétaux en Agriculture de Conservation des Sols. Témoignages de l’expérience de 2 frères agriculteurs de l’APAD 62, situé dans le Marquenterre, Somme (80) dans la région des Hauts-de-France. 🐑 Présentation du troupeau. 🐑 Comment gérez-vous le pâturage des couverts sur la ferme ? 🐑 Quel est l’impact du troupeau sur le sol ? 🐑 Quelle est la place des couverts au niveau de la rentabilité ? 🐑 Quels conseils donneriez-vous à quelqu’un qui souhaite se lancer dans cette pratique ?
La présence d’un troupeau d’ovin dans les parcelles en SCV accélèrent la décomposition des plantes et augmentent la vie bactérienne positivement …!!
L’ ACS expliqué sur l’excellent site triple performance https://wiki.tripleperformance.fr/wiki/Triple_Performance
L’agriculture de conservation des sols (ACS) est un ensemble de techniques culturales destinées à maintenir et améliorer le potentiel agronomique des sols, tout en conservant une production régulière et performante sur les plans technique et économique.
99,99 % Des Pesticides Que Nous Consommons Sont Fabriqués Par Des Plantes
Autant ce que l’on appelle pesticides sont retrouvés NATURELLEMENT jusque dans notre alimentation…..Autant dans la nature vous ne trouverez aucun exemple de travail mécanique de sol comme l’agriculture conventionnelle le pratique . Les plantes n’ont pas besoin de travail mécanique de sol pour se développer …..Mais elles ont besoin de fabriquer des produits pour leurs défenses auprès des prédateurs et des maladies
Quand Bruce Ames parle de toxicité, il est temps d’écouter (1) . Ames est l’inventeur du très important test d’Ames pour la mutagénicité, qui mesure les dommages causés à l’ADN par un produit chimique donné. Le test d’Ames constitue un obstacle essentiel dans le monde de la recherche sur la découverte de médicaments. Même si un test Ames positif ne constitue pas une preuve de facto qu’un produit chimique sera cancérigène chez l’homme, il s’agit d’un signal d’alarme géant dans le développement de médicaments. De nombreux médicaments prometteurs ont rencontré leur fabricant simplement grâce à un test d’Ames positif.
BONNE CHANCE POUR ÉVITER LES PESTICIDES
Étant donné que les pesticides et les herbicides font régulièrement la une des journaux, dernièrement à cause de la « guerre du glyphosate », (2) j’ai pensé qu’il pourrait être intéressant d’examiner un article de synthèse qu’Ames et ses collègues ont écrit il y a près de trois décennies dans les Actes de l’Académie nationale des sciences. (PNAS) . Si vous vivez dans l’illusion que vous pouvez vous lancer dans des aliments entiers et payer trop cher pour un tas de produits biologiques sans pesticides, cet article vous détrompera de cette erreur. Si vous ne voulez pas consommer de pesticides, alors vous feriez mieux d’arrêter de manger, car vous en consommez à chaque bouchée. Il y en a beaucoup. Selon Ames :
99,99 % (en poids) des pesticides consommés par le public américain sont fabriqués par les plantes elles-mêmes comme mécanisme de défense.
Les pesticides synthétiques et naturels sont également susceptibles d’être cancérigènes.
(Il est peu probable que vous voyiez cette information publiée dans Whole Foods.)
Ames écrit (c’est moi qui souligne) :
« L’examen toxicologique des produits chimiques synthétiques tels que les pesticides et les polluants industriels, sans examen similaire des produits chimiques présents dans la nature à des fins de comparaison, a généré un déséquilibre à la fois dans les données et dans la perception des dangers potentiels pour les humains . »
Sans blague. À l’American Council, nous crions cela depuis de nombreuses années, pour ensuite être traités de « complices de l’industrie » chaque fois que nous osons qualifier un produit chimique de sûr. Non seulement nous ne sommes pas des complices (3) , mais nous avons toujours eu raison sur un point clé. La distinction entre produits chimiques « naturels » et « artificiels » n’a aucun sens – ce que beaucoup d’Américains ne connaissent pas. L’origine d’un produit chimique n’a pas d’importance. Votre corps ne peut pas dire ce qui est naturel ou synthétique – seulement les propriétés de la matière chimique.
Il est difficile de blâmer les consommateurs ; l’industrie alimentaire biologique et les charlatans d’Internet ont fait un travail magistral en créant un récit selon lequel nous sommes tous empoisonnés par d’infimes quantités de milliers de produits chimiques fabriqués par l’homme, et que le moyen d’éviter cela est d’acheter des produits dits « naturels ». produits car ils ne contiennent pas de produits chimiques. Mais le récit est complètement faux. Excellente commercialisation. Terrible science. Voyons comment Ames est parvenu à ses conclusions.
MODÈLES DE CANCÉROGÉNICITÉ DU RAT ET DE LA SOURIS – DE NOMBREUSES LIMITES
Toutes les données de cancérogénicité contenues dans l’article du PNAS proviennent de modèles de cancer chez le rat et la souris, qui sont notoirement peu fiables pour prédire les cancers humains, mais ce sont les meilleurs que nous ayons. Ames et ses collègues de l’UC Berkeley ont parcouru la littérature à la recherche d’études sur les aliments contenant des pesticides naturels connus. Les chiffres sont énormes. Par exemple, rien que dans le chou, 49 pesticides et leurs métabolites ont été détectés. Parmi ceux-ci, deux produits chimiques, l’acide chlorogénique et l’isothiocyanate d’allyle, ont provoqué des tumeurs chez le rat, mais pas chez la souris (4) . Ames estime que les Américains consomment entre 5 000 et 10 000 pesticides naturels différents.
LES CANCÉRIGÈNES SONT PARTOUT
La revue a également identifié des études dans lesquelles des pesticides d’origine végétale avaient été testés (à fortes doses) (5) pour voir s’ils provoquaient le cancer chez les rongeurs. Cinquante-deux de ces pesticides, tous couramment présents dans divers aliments, ont été évalués de cette manière ; 27 se sont révélés cancérigènes. Plus de la moitié.
« Les pesticides naturels constituent un sous-ensemble important de produits chimiques naturels. Les plantes produisent des toxines pour se protéger contre les champignons, les insectes et les prédateurs animaux. »
Bruce Ames, et. Al.
Les plantes n’existent pas pour servir les humains ; ils sont ici parce qu’ils ont survécu et se sont reproduits. Pour ce faire, ils ont développé la capacité de synthétiser des produits chimiques pour les protéger des prédateurs. Malgré les protestations constantes contre les pesticides, nous n’aurions rien à manger si les plantes ne produisaient pas celles dont elles ont besoin pour se défendre.
ET NOUS EN MANGERONS BEAUCOUP
Notre obsession idiote d’éviter d’infimes quantités de résidus de pesticides sur une pomme semble encore plus idiote lorsque nous examinons les quantités relatives de pesticides naturels et synthétiques que nous consommons. Lorsque la FDA a analysé les aliments pour détecter la présence de produits chimiques synthétiques importants (probablement présents dans l’environnement), 105 résidus chimiques différents ont été détectés dans les aliments (6) . La somme totale de ces 105 produits chimiques (combinés) a été estimée à environ 0,09 mg par personne et par jour, dont environ la moitié sont cancérigènes. En revanche, nous consommons quotidiennement environ 1,5 g (1 500 mg) de pesticides naturels.
Nous concluons également qu’aux faibles doses de la plupart des expositions humaines, les risques comparatifs des résidus de pesticides synthétiques sont insignifiants .
MAIS NOUS NE SOMMES PAS TOUS MORTS
Nous consommons régulièrement des milliers de pesticides, la plupart fabriqués par les plantes, et une quantité bien moindre de résidus de pesticides appliqués sur les cultures. Et pourtant, nous sommes toujours là. La raison est évidente, et c’est la même raison pour laquelle il n’y a pas de quoi s’inquiéter des infimes quantités de croque-mitaines environnementaux omniprésents comme le BPA, les phtalates et les parabènes : la dose. Les produits chimiques cancérigènes, qu’ils soient d’origine végétale ou synthétiques, sont traités par le foie et excrétés. C’est le travail du foie, et il le fait très bien. Sinon, il n’y aurait pas besoin de pesticides de synthèse car le chou nous aurait déjà planté en terre.
REMARQUES:
(1) Bruce Ames, l’un des fondateurs de l’American Council, n’a pas remporté le prix Nobel pour son invention. Cela me laisse perplexe.
(2) Même si cela n’a rien de drôle, il y a eu beaucoup d’affaires amusantes concernant le glyphosate ces derniers temps, y compris des fraudes. En parlant de fraude, voyez le démantèlement brutal de la corruption du CIRC par mon collègue Alex Berezow . Le Dr Berezow a également écrit ici sur les pesticides à base de plantes .
(3) Des compères ? À peine. À l’ACSH, nous gardons un mur entre la science et la collecte de fonds pour éviter ne serait-ce que l’apparence d’un conflit d’intérêts. À l’heure actuelle, c’est un mur inutile. On m’a dit que 97 % de notre financement provenait de donateurs individuels. Je ne sais pas d’où viennent les 3% restants.
(4) Le statut réglementaire actuel de l’isothiocyanate d’allyle n’est pas cohérent avec les conclusions d’Ames. Le produit chimique n’apparaît pas sur la liste de la proposition 65 de Californie et fait partie du groupe 3 du CIRC – non classifiable quant à sa cancérogénicité pour l’homme.
(4) Cet écart est expliqué par le Programme national de toxicologie car les souris ne reçoivent pas la dose maximale tolérée des deux produits chimiques alors que les rats la reçoivent.
(5) Je ne plaisante pas sur les fortes doses. Environ la moitié de ces tests ont été réalisés à la dose maximale tolérée (DMT) – la dose à laquelle les animaux commencent à mourir si elle est dépassée. On ne sait pas si cela a à voir avec le fait que les humains consomment de petites quantités de ce produit chimique – une autre limite des tests de cancérogénicité sur les rongeurs.
(6) Conseil national de recherches, Conseil de l’agriculture (1987) Réglementation des pesticides dans les aliments (National Academy Press, Washington, DC).
A mon humble avis, le seul truc qui peut avoir l’ambition de résoudre un panel assez large de problématiques actuelles de cette planète, s’appelle la PHOTOSYNTHÈSE , la photosynthèse qui fait la force de la Nature est la fonction inimitable ( à énergie gratuite) qui enclenche tous les processus qui intéressent toute la vie sur cette Terre …..On ne parle pas assez de cette photosynthèse ….On l’a plutôt sacrement mise à mal depuis quelques temps …..On doit peut-être remettre « de la photosynthèse dans les cerveaux »
Si l’homme était si intelligent, au lieu d’avoir une planète avec un climat détraqué, on devrait avoir un paradis naturel en évolution constante……Uniquement si on avait un peu compris la Nature ….au lieu de la détruire , il faut la développer , la booster, et ce qui est terrible , c’est qu’elle peut le faire seule, il suffit de la laisser tranquille …..Elle sait ce qu’elle doit faire faire pour être performante …..Elle faisait ça tranquille depuis très, très longtemps……La Nature est d’une puissance magnifique
Article présentant les différentes étapes de la photosynthèse et des expériences réalisables en classe permettant de montrer le déroulement de celle-ci. Analyse de résultats expérimentaux et de démonstrations célèbres : expériences d’Emerson, d’Engelman, de Ruben et Kamen…
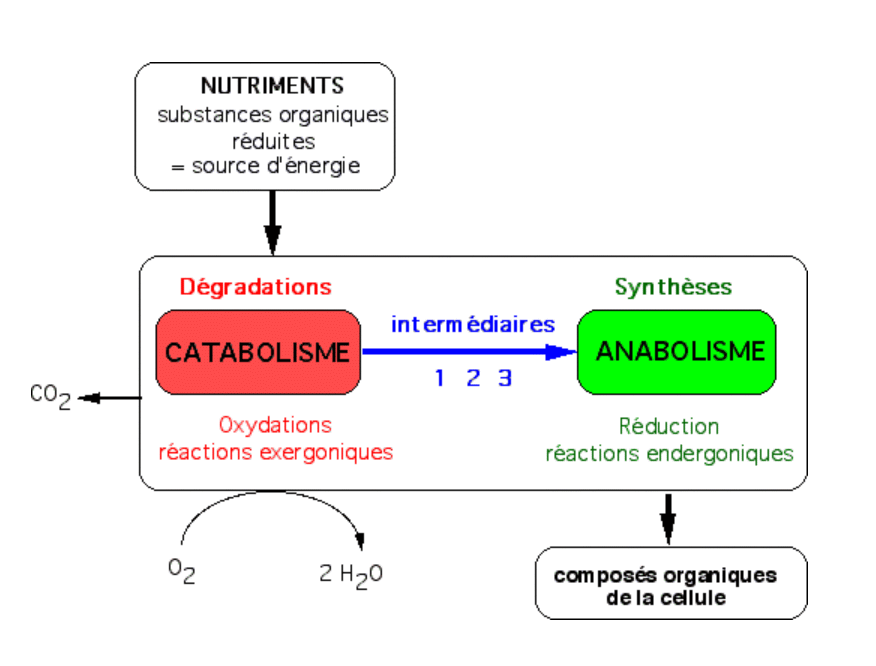
1. Introduction
Les végétaux, organismes photoautotrophes, sont capables d’utiliser l’énergie lumineuse pour réaliser la synthèse de molécules organiques, à partir de composés minéraux. L’ensemble de ces réactions est regroupé sous le terme de photosynthèse. La photosynthèse est réalisée par des organismes autotrophes au carbone, grâce à des pigments particuliers, et peut être découpée en deux groupes de réactions.
Le dossier aborde de manière succincte ces généralités sur la photosynthèse. Il s’agit essentiellement d’une version « abrégée » de l’ensemble de documents présents sur le site Biologie et Multimédia qui reprennent l’essentiel du module « Biologie et Physiologie végétales » de 2e année de l’Université Paris VI. Ce dossier « abrégé » reste donc bien évidemment incomplet. À tout moment, il est possible d’accéder aux documents complets, par les liens signalés.
2. Les organismes autotrophes au carbone
2.1. Autotrophie et hétérotrophie
Les êtres vivants sont composés d’eau et de sels minéraux, ainsi que de substances organiques. Ces dernières comportent glucides, lipides, protéines, acides nucléiques, etc. Or les composés organiques sont continuellement renouvelés (par dégradation et synthèse). Ce fonctionnement des êtres vivants nécessite des échanges constants de matière et d’énergie avec le milieu extérieur.
On peut ainsi distinguer différents types d’organismes en fonction de leurs besoins et de la source d’énergie utilisée.
Les organismes hétérotrophes : ils sont incapables d’effectuer eux-mêmes les synthèses de leurs constituants à partir d’élément minéraux. Ils sont en général chimiotrophes, c’est-à-dire utilisant comme source d’énergie l’énergie chimique récupérée au cours de l’oxydation des composés organiques réduits présents dans leur alimentation.
Les organismes autotrophes : ils sont capables d’utiliser des éléments inorganiques pour synthétiser leurs propres constituants organiques. Ils sont en général phototrophes, c’est-à-dire capables d’utiliser l’énergie lumineuse et de convertir cette énergie en étapes chimiques.
Il s’agit des animaux, des champignons, et de certains procaryotes (la bactérie E. coli par exemple). Ces organismes utilisent des substances organiques à la fois comme source d’énergie et comme source de pouvoir réducteur.
Figure 1 – Schéma général du métabolisme d’une cellule hétérotrophe / chimiotropheLes intermédiaires sont : 1 = ATP, intermédiaire énergétique ; 2 = NADH ou NADPH, coenzymes d’oxydo-réduction ; 3 = molécules du métabolisme intermédiaire (pyruvate, malate, acétylCoA, etc.).Auteur(s)/Autrice(s) : Roger Prat, François MoreauLicence : Pas de licence spécifique (droits par défaut)
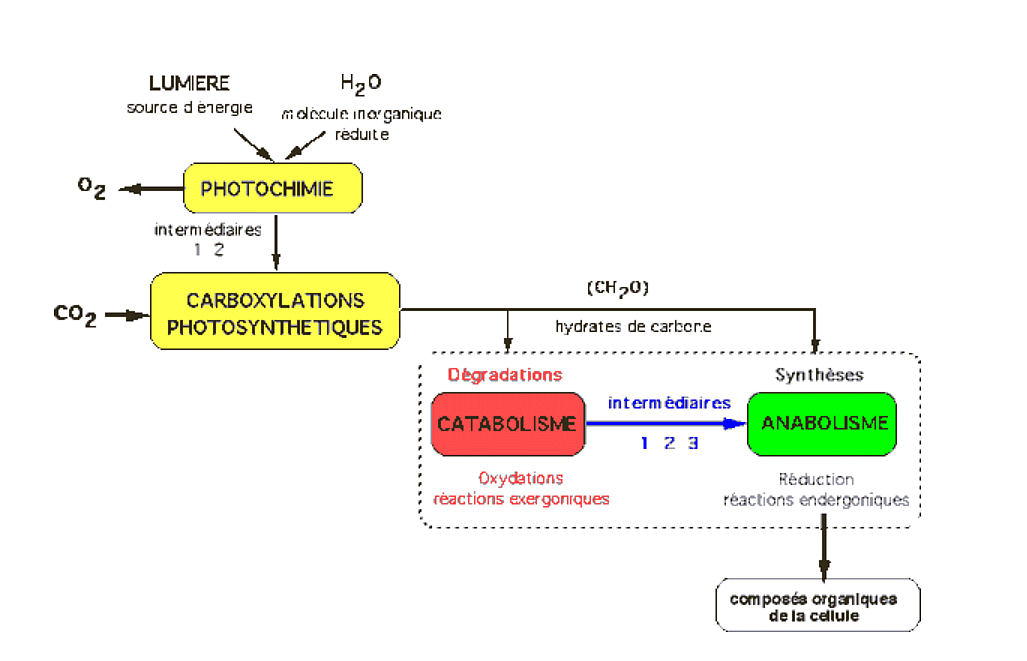
Il s’agit des végétaux chlorophylliens et de certains procaryotes. Ces organismes utilisent la lumière comme source d’énergie et l’eau comme pouvoir réducteur.
Figure 2 – Schéma général du métabolisme d’une cellule autotrophe / phototropheLes intermédiaires sont : 1 = ATP, intermédiaire énergétique ; 2 = NADH ou NADPH, coenzymes d’oxydo-réduction ; 3 = molécules du métabolisme intermédiaire (pyruvate, malate, acétylCoA, etc.).Auteur(s)/Autrice(s) : Roger Prat, François MoreauLicence : Pas de licence spécifique (droits par défaut)
Toutefois, le fait qu’un organisme est autotrophe n’implique pas que toutes ses cellules soient autotrophes. Ainsi, si l’on prend l’exemple des trachéophytes (plantes vascularisées, ce qui inclus les plantes à fleurs), on peut noter que dans leur cas l’appareil aérien est autotrophe, mais que l’appareil racinaire est lui hétérotrophe (de même que l’embryon et la plantule).
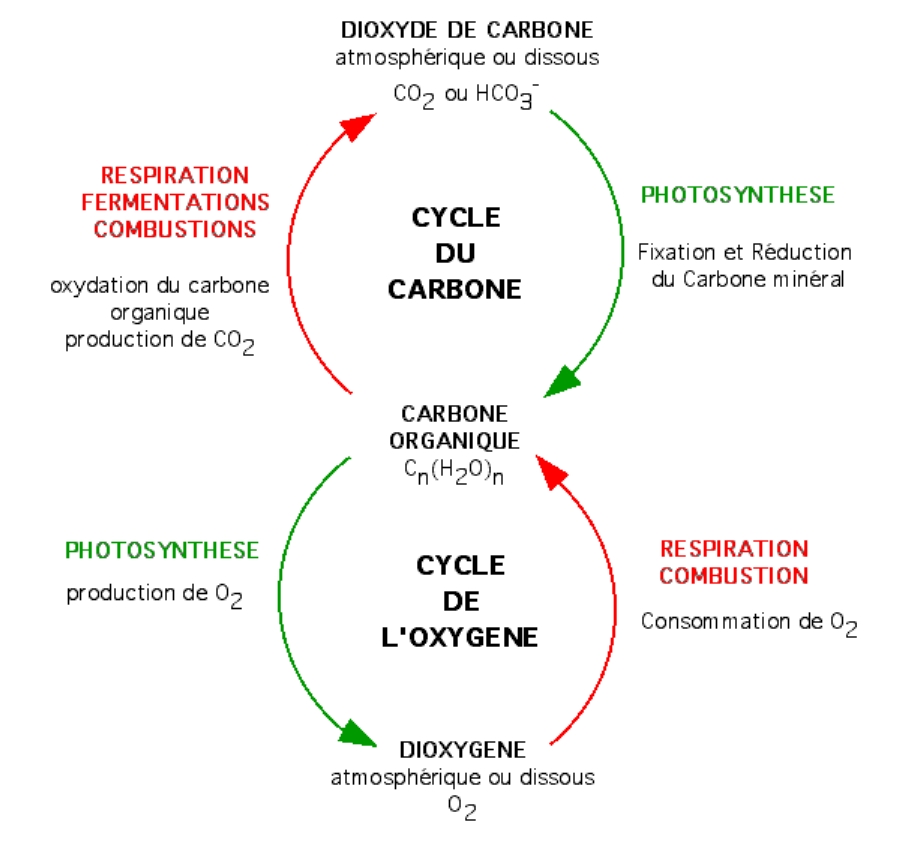
Figure 3 – Cycle du carbone et cycle de l’oxygèneLa photosynthèse des végétaux chlorophylliens est responsable de la fixation et de la réduction de CO2, ainsi que de la libération d’O2. À l’inverse, la fonction respiratoire des organes et organismes non chlorophylliens est responsable de l’oxydation des composés organiques (consommation d’O2, libération de CO2). Il en résulte un cycle pour le carbone et un cycle pour l’oxygène qui sont antiparallèles.Auteur(s)/Autrice(s) : Roger Prat, François MoreauLicence : Pas de licence spécifique (droits par défaut)
Chez les plantes terrestres (Embryophytes, anciennement appelées cormophytes), la photosynthèse se réalise dans les chloroplastes des parenchymes chlorophylliens des organes chlorophylliens. Ces organes sont les feuilles, plus rarement les tiges. Chez les algues, les cellules chlorophylliennes sont localisées dans l’ensemble du thalle.
Nous nous limitons ici à l’exemple des Angiospermes. Une étude expérimentale (par exemple basée sur la présence d’amidon, stocké temporairement lors de la photosynthèse) permet de mettre en évidence la localisation de la photosynthèse, aussi bien au niveau de l’organisme dans son entier qu’au sein de la cellule elle-même (voir à ce sujet le document sur cette mise en évidence expérimentale)
3.1. Localisation au sein des feuilles
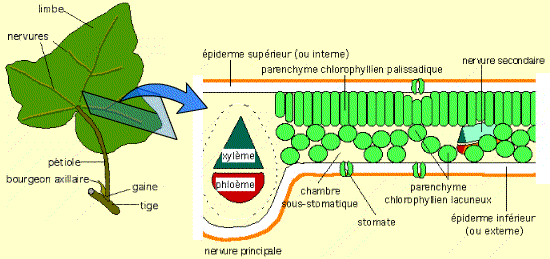
Chez les Angiospermes, la photosynthèse est essentiellement localisée au niveau de la feuille. Cet organe aplati, en relation étroite avec la tige, possède une morphologie lui permettant de présenter une grande surface vis-à-vis de l’environnement.
Figure 4 – Structure schématique d’une feuille d’Angiosperme dicotylédoneLa nervure médiane, très en relief comme chez beaucoup de dicotylédones, contient principalement des tissus conducteurs de la sève brute (xylème) et de la sève élaborée (phloème). Ces tissus sont protégés par des tissus de soutien. De part et d’autre de cette nervure, le limbe est formé par du parenchyme palissadique (face supérieure) et du parenchyme lacuneux (face inférieure). La feuille est protégée des pertes d’eau par deux épidermes, recouverts d’une cuticule imperméable. Les échanges de gaz sont assurés par les stomates.Auteur(s)/Autrice(s) : Roger Prat, François MoreauLicence : Pas de licence spécifique (droits par défaut)
La plupart des feuilles d’Angiospermes dicotylédones présente un parenchyme chlorophyllien palissadique à la face supérieure : c’est à son niveau que se déroule la photosynthèse. Ce tissu est en relation aussi bien avec l’extérieur (par les stomates) qu’avec l’intérieur de la plante (par les tissus conducteurs des nervures).
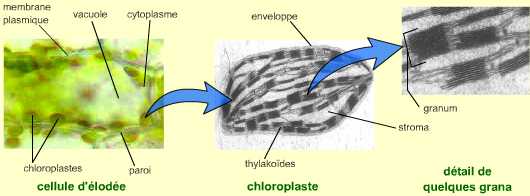
Au sein des cellules chlorophylliennes, la photosynthèse se déroule dans les chloroplastes. Ces organites de grande taille (environ 10 micromètres de long) possèdent une enveloppe composée d’une double membrane, et un système endomembranaire formant des saccules : les thylakoïdes. La conversion de l’énergie lumineuse en énergie de liaison chimique et en pouvoir réducteur se réalise au niveau des membranes des thylakoïdes. La réduction du carbone inorganique (CO2) en carbone organique a lieu dans le stroma du chloroplaste. Cette matière organique synthétisée peut être stockée temporairement sous la forme de grains d’amidon.
Figure 5 – Photographies d’une cellule chlorophyllienne (d’élodée du Canada) et d’un chloroplasteDans la cellule végétale, les chloroplastes sont disposés dans le cytoplasme périphérique de la vacuole. Voir le document complet pour un schéma explicatif. Le chloroplaste est observé au microscope électronique à transmission. On note deux types de thylakoïdes : les thylakoïdes granaires qui s’assemblent en « piles » de saccules (les grana), et les thylakoïdes intergranaires.Auteur(s)/Autrice(s) : Roger Prat, François MoreauLicence : Pas de licence spécifique (droits par défaut)
Les chloroplastes peuvent être observés dans les conditions naturelles (« in situ »). Mais il est parfois nécessaire de les isoler, afin de réaliser une étude plus précise de leur nature et de leur fonctionnement. Pour cela, on procède à l’éclatement des cellules végétales, puis à l’isolement et à la purification des chloroplastes intacts par plusieurs centrifugations successives.
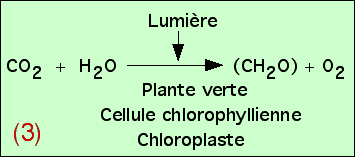
Diverses expériences permettent d’aboutir à une équation globale, résumant les mécanismes de la photosynthèse. Nous revenons ici sur quelques expériences permettant d’en démontrer les différents éléments, et donc de construire progressivement cette équation.
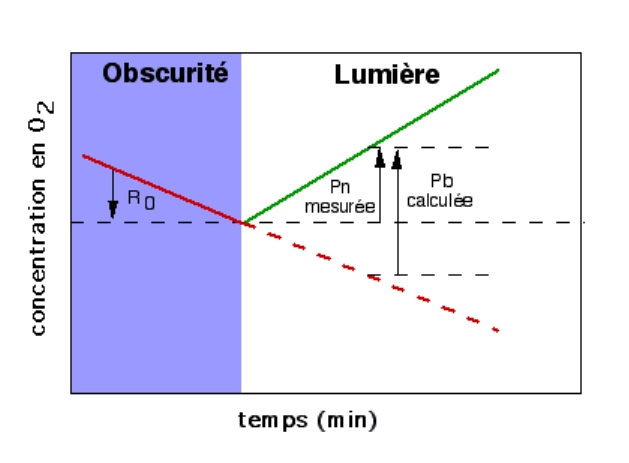
4.1. Production de dioxygène, utilisation de dioxyde de carbone
On peut tout d’abord chercher si certains échanges gazeux se réalisent chez les plantes chlorophylliennes, en présence de lumière. On utilisera pour cela une plante aquatique, l’élodée du Canada, et comme source de CO2, de l’hydrogénocarbonate de sodium. Celui-ci, soluble dans l’eau est absorbé par la plante et converti en CO2 grâce à une anhydrase carbonique selon la réaction :
Figure 6 – Expérience de dégagement de dioxygène par une élodée à la lumièreLes trois expériences sont réalisées dans : (a) de l’eau distillée ; (b) de l’eau du robinet ; (c) de l’eau additionnée d’hydrogénocarbonate à 1 %. C’est en (c) que la production d’oxygène est la plus importante.Auteur(s)/Autrice(s) : Roger Prat, François MoreauLicence : Pas de licence spécifique (droits par défaut)Figure 7 – Équation bilan de la photosynthèse : équation (1)L’expérience précédente montre qu’à la lumière, une plante verte produit de l’O2 si du CO2 lui est fourni. Cette constatation n’implique aucune relation chimique entre le CO2 et l’O2.Auteur(s)/Autrice(s) : Roger Prat, François MoreauLicence : Pas de licence spécifique (droits par défaut)
Ces expériences permettent donc de démontrer qu’en présence de lumière, les végétaux chlorophylliens consomment du CO2 et libèrent du O2. Toutefois, ces expériences seules ne nous permettent pas d’expliquer ce que permettent ces échanges gazeux pour la plante.
4.2. Production de glucides
Dans un deuxième temps, on recherche si l’exposition à la lumière a des conséquences sur la matière organique (et plus particulièrement glucidique) présente au sein du végétal. Des expériences utilisant des isotopes radioactifs démontrent ainsi que l’énergie lumineuse permet, indirectement, la synthèse de glucides simples.
Toutefois, il est difficile de caractériser ces glucides simples produits par la photosynthèse dans des expériences utilisant du matériel simple. Il est possible par contre de caractériser l’amidon (un polymère de glucose mis en réserve lorsque la photosynthèse est très active). Cette caractérisation se réalise avec le lugol, un réactif spécifique de l’amidon.
On peut ainsi observer la présence d’amidon au sein des chloroplastes de cellules de feuille d’élodée mises à la lumière.
Figure 8 – Observation d’une feuille d’élodée exposée à la lumièreUne feuille d’élodée est placée dans une eau enrichie en hydrogénocarbonate et éclairée plusieurs heures. A gauche : cellules observées sans coloration, chloroplastes naturellement verts. A droite : après traitement par le lugol, des grains d’amidon de couleur sombre sont visibles dans les chloroplastes.Auteur(s)/Autrice(s) : Roger Prat, François MoreauLicence : Pas de licence spécifique (droits par défaut)
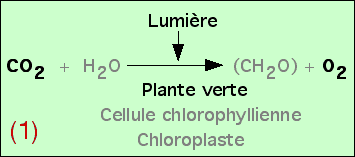
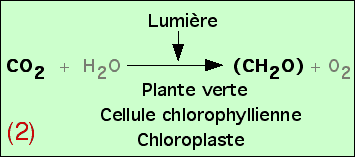
On obtient ainsi l’équation bilan de la photosynthèse. Afin d’obtenir un équilibre chimique de cette réaction, on rajoute H2O, mais sans que les expériences présentées ici aient permis de démontrer son utilisation réelle.
Des études plus précises peuvent être réalisées, afin de mieux comprendre les relations entre les atomes des molécules figurées dans cette équation bilan. Si le devenir du carbone du CO2 ne pose pas de problème (il est incorporé dans les glucides synthétisés), l’origine de l’oxygène de l’O2 pourrait se trouver soit au niveau du CO2, soit au niveau de l’eau H2O. En réalité, il apparaît que c’est l’oxygène de l’eau qui est libéré, au cours d’une réaction d’oxydo-réduction. Ceci permet de préciser alors l’équation bilan de la photosynthèse.
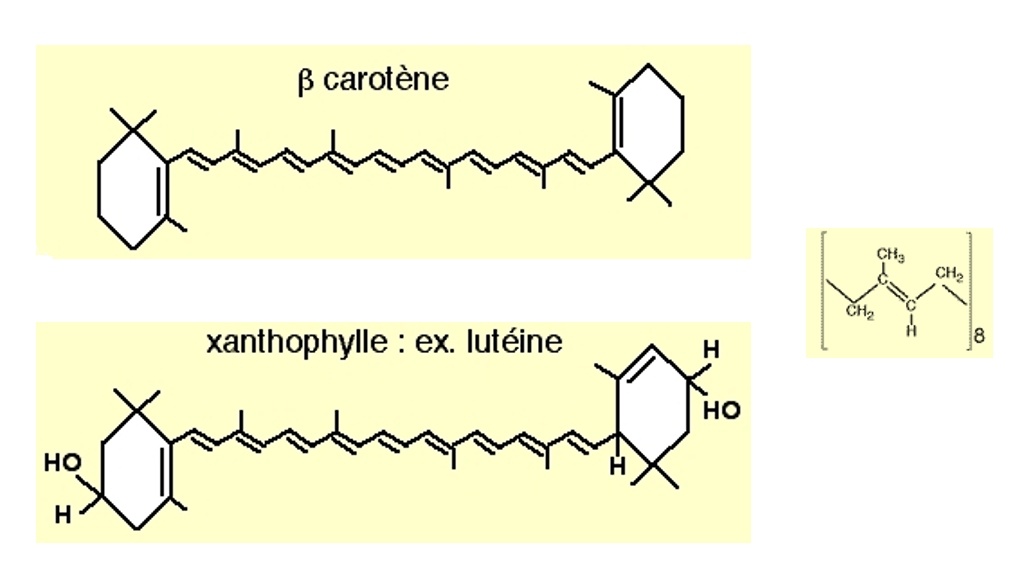
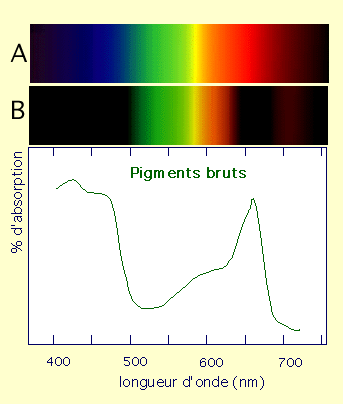
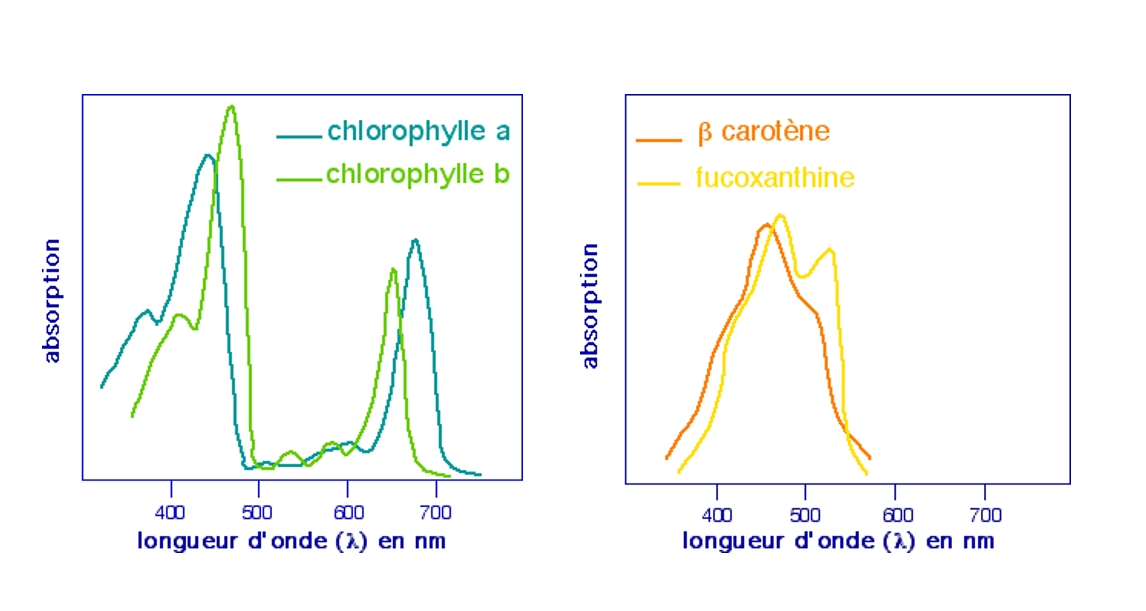
La réalisation de la photosynthèse par les chloroplastes des végétaux met en jeu un ensemble de molécules particulières, nommées pigments photosynthétiques. Le terme de « pigment » correspond au fait que ces molécules sont colorées, de part leur capacité à capter certaines radiations lumineuses. Ces pigments sont de trois types :
les chlorophylles, présentes chez tous les végétaux autotrophes au carbone ;
les caroténoïdes, présents chez tous les végétaux autotrophes au carbone ;
les phycobilines, présentes exclusivement chez les algues et les cyanobactéries.
On peut assez facilement extraire et séparer ces différents pigments.
Les chlorophylles sont constituées d’un noyau tétrapyrrolique avec un magnésium en son centre, et estérifié avec un alcool à très longue chaîne en C20 (le phytol). Dans la membrane des thylakoïdes, les chlorophylles sont associées à des protéines et forment des complexes protéines – pigments.
Figure 11 – Formules des chlorophylles a et bLes chlorophylles diffèrent par les substituants des groupements pyrroles. Le phytol n’est pas détaillé ici. Légende : I, II, III, IV = groupements pyrroles et V = cycle supplémentaire.Auteur(s)/Autrice(s) : Roger Prat, François MoreauLicence : Pas de licence spécifique (droits par défaut)
Télécharger le fichier PDB de la chlorophylle a pour une visualisation avec rasmol / rastop : chloa.pdb
Les caroténoïdes sont des molécules constituées de 40 carbones, avec deux extrémités cyclisées reliées par une longue chaîne de 8 unités isoprènes.
Figure 12 – Formule de deux caroténoïdesLe β-carotène est un exemple de carotène, et la lutéine un exemple de xanthophylle. À droite est représentée une unité isoprène.Auteur(s)/Autrice(s) : Roger Prat, François MoreauLicence : Pas de licence spécifique (droits par défaut)
Télécharger le fichier PDB du βcarotène pour une visualisation avec rasmol / rastop : bcarotene.pdb
Les phycobilines sont composées d’un noyau tétrapyrrolique ouvert, associé à une protéine. On les trouve au sein des photosystèmes de certaines algues, et de bactéries photosynthétiques telles que les cyanobactéries.
Figure 13 – Formule d’une phycobilineL’exemple présenté ici est la phycocyanobiline, représentée sans la protéine qui l’accompagne normalement.Auteur(s)/Autrice(s) : Roger Prat, François MoreauLicence : Pas de licence spécifique (droits par défaut)
Télécharger le fichier PDB de la phycocyanobiline pour une visualisation avec rasmol / rastop : phycocyanobiline.pdb
5.2. Spectres d’absorption
Les chlorophylles et les caroténoïdes absorbent certaines radiations dites actives pour la photosynthèse, dans la gamme de longueurs d’onde visibles comprises entre 500 et 700 nm.
À partir d’une solution de pigments, on peut donc mesurer les caractéristiques d’absorption de la lumière en réalisant un spectre d’absorption à l’aide d’un spectrophotomètre UV-visible classique, qui permet de mesurer l’absorption (A) en fonction de la longueur d’onde (l).
Figure 14 – Spectre d’absorption des pigments bruts extraits à partir d’une feuilleA : spectre lumineux en absence de pigments. B : spectre lumineux en présence de pigments. On note que l’absorption maximale se réalise dans le bleu et dans le rouge.Auteur(s)/Autrice(s) : Roger Prat, François MoreauLicence : Pas de licence spécifique (droits par défaut)
Un tel spectre global ne permet pas de reconnaître la part qui revient à chaque pigment. Pour cela, il faut travailler sur des solutions de pigments séparés et purifiés.
Figure 15 – Spectres d’absorption des chlorophylles, du carotène et de la fucoxanthineA gauche : spectres d’absorption des chlorophylles a et b. A droite : spectres d’absorption du bêta-carotène et de la fucoxanthineAuteur(s)/Autrice(s) : Roger Prat, François MoreauLicence : Pas de licence spécifique (droits par défaut)
Les chlorophylles sont des pigments. De ce fait, ces molécules (comme les autres pigments photosynthétiques) peuvent être excitées par les radiations lumineuses. Cette excitation est due à la présence de liaisons conjuguées (et donc d’électrons délocalisés) : l’arrivée d’un photon fait passer un électron délocalisé d’un état fondamental (non excité) à un état excité. Chez la chlorophylle, il existe deux états excités : un état supérieur (Sa) et un état inférieur (Sb), selon l’énergie du photon excitateur.
La chlorophylle, une fois excitée, retourne à son état fondamental, plus stable thermodynamiquement. Ceci peut se faire de plusieurs manières, et en particulier en :
émettant de la lumière (c’est la fluorescence constatée dans une solution de chlorophylle) ;
transférant son énergie à une molécule très proche (c’est la résonance, qui permet aux pigments de l’antenne collectrice des photosystèmes de transférer l’énergie lumineuse de molécule en molécule jusqu’à une chlorophylle piège) ;
perdant un électron (c’est la photochimie, qui permet à la molécule de chlorophylle piège du photosystème de réduire un accepteur d’électron, et ainsi de permettre la réalisation de la chaîne photosynthétique).
Les spectres d’absorption des pigments sont uniquement liés à leur capacité à capter des photons de certaines longueurs d’onde. Le spectre d’action consiste à quantifier l’activité que l’on cherche à corréler à ces pigments (ici l’activité photosynthétique), en fonction des longueurs d’onde incidentes. Les spectres d’action ainsi réalisés suivent globalement les spectres d’absorption des végétaux chlorophylliens, ce qui confirme que c’est bien cette capacité à capter les photons qui permet la réalisation de la photosynthèse.
La photosynthèse est influencée par les facteurs de l’environnement : la lumière (source d’énergie), le CO2 (source de carbone) et la température (qui affecte l’ensemble des réactions biochimiques).
La photosynthèse est un processus complexe qui fait intervenir de nombreuses étapes qui sont affectées de manière différente par les facteurs de l’environnement. De ce fait, les facteurs externes agissent indépendamment les uns des autres et le phénomène global obéit à la loi dite des « facteurs limitants » que l’on peut énoncer de la façon suivante : lorsqu’un processus est contrôlé par plusieurs facteurs agissant indépendamment, son intensité est limitée par le facteur qui présente la valeur minimum. Le facteur est alors limitant et la vitesse du processus est proportionnelle à la valeur de ce facteur.
6.1. Mesure de la photosynthèse
Pour pouvoir étudier les facteurs externes influant sur la photosynthèse, encore faut-il être capable de mesurer celle-ci. Dans cette optique, un certain nombre de paramètres peuvent être pris en compte, et en particulier l’incorporation du carbone dans les molécules organiques, l’évolution de la concentration en CO2, ou encore l’évolution de la concentration en oxygène.
Une solution simple et quantitative est l’utilisation d’une électrode à oxygène pour mesurer l’évolution de la concentration en oxygène. Ainsi, on observe à la lumière un dégagement d’oxygène. La mesure de ce dégagement correspond à la photosynthèse nette (Pn). En effet, la plante, dans le même temps, réalise la respiration cellulaire, et consomme ainsi de l’oxygène, ce qui fausse cette mesure… La solution est alors de mesurer la consommation d’oxygène à l’obscurité, qui correspond à la respiration (Ro). On obtient alors la valeur de la photosynthèse brute (Pb) par la formule suivante : Pb = Pn – Ro
Figure 18 – Évolution de la concentration en dioxygène à l’obscurité et à la lumière lors de la photosynthèsePb = Pn – RoPb = photosynthèse brute ; Pn = photosynthèse nette ; Ro = respirationAuteur(s)/Autrice(s) : Roger Prat, François MoreauLicence : Pas de licence spécifique (droits par défaut)
La photosynthèse se réalise en présence de lumière. Il est possible de quantifier ce phénomène, en éclairant des plantes avec une source lumineuse permettant de réaliser une gamme d’intensités (flux de photons) déterminées.
On obtient ainsi des courbes biphasiques, permettant de déterminer plusieurs paramètres :
L’éclairement saturant ou optimal (IS) : c’est l’éclairement pour lequel la courbe atteint un plateau. Au-delà, la capacité d’absorption des photons dépasse la capacité de leur utilisation. Les réactions d’assimilation du CO2 deviennent limitantes et la photosynthèse présente une intensité maximale.
Le point de compensation pour la lumière (IC) : c’est la valeur de l’éclairement pour laquelle la photosynthèse nette est nulle ; la photosynthèse compense juste la respiration.
Le rendement de l’absorption des photons (ou rendement quantique foliaire Phi Ф) c’est la pente (coefficient directeur) de la partie linéaire initiale de la courbe. Dans cette gamme d’éclairement, la lumière est limitante.
Il est aussi possible d’étudier l’influence qualitative de la lumière, en réalisant le spectre d’action de la lumière sur le végétal étudié. On peut ainsi s’apercevoir que toutes les radiations lumineuses ne sont pas aussi efficaces pour la photosynthèse.
6.3. Comparaison de la photosynthèse de plantes de lumière et de plantes d’ombre
Figure 20 – Courbes de saturation de la photosynthèse en fonction de la densité du flux de photons chez une plante de lumière et une plante d’ombreLes autres facteurs (concentration en CO2 atmosphérique, température 25 °C) sont maintenus constants. IC : intensité de compensation ; IS : intensité saturante ; Ф : rendement quantique foliaire. En bleu : plantes d’ombre ; en rouge : plantes de lumière.Auteur(s)/Autrice(s) : Roger Prat, François MoreauLicence : Pas de licence spécifique (droits par défaut)
Quand on compare le comportement de ces deux types de plantes on constate que :
ICO (ombre) est inférieure à ICL (lumière) ;
ФO (ombre) est supérieur à ФL (lumière) ;
ISO (ombre) est inférieure à ISL (lumière).
En d’autres termes, les plantes d’ombre présentent une intensité photosynthétique optimale et une intensité de compensation plus faible, mais une efficacité dans l’absorption des photons plus élevée (plantes des sous-bois). Inversement, les plantes de lumière sont moins efficaces dans la capture des photons, mais elles fixent davantage de CO2 (ex : plantes cultivées).
6.4. Influence de la concentration en CO2
Les plantes aériennes assimilent le CO2 atmosphérique (0,035 % de CO2) tandis que les plantes aquatiques absorbent soit le CO2 dissous (concentration faible : environ 10 µM à pH 7), soit les ions bicarbonate HCO3– (concentrations élevées : de l’ordre du mM, mais variable en fonction du pH), qui sont ensuite convertis en CO2 grâce à la réaction catalysée par l’anhydrase carbonique. La quantité de CO2 disponible est limitante dans des conditions d’éclairement moyen. Par conséquent, une augmentation de la photosynthèse est observée lorsqu’on augmente la concentration de CO2.
Figure 21 – Influence de la concentration en CO2 de l’air sur la consommation en CO2 d’une plante verteLa courbe présente une première partie pseudo-linéaire pour laquelle le CO2 est limitant, et une seconde partie qui correspond à un plateau pour lequel l’éclairement est devenu limitant et la photosynthèse maximale, dans ces conditions.Auteur(s)/Autrice(s) : Roger Prat, François MoreauLicence : Pas de licence spécifique (droits par défaut)
L’optimum de température des plantes varie en fonction de leur origine. Ainsi, les plantes des régions tempérées ont un maximum qui se situe entre 15 °C et 25 °C, avec une limite de tolérance au froid vers – 2 °C à 0 °C et de tolérance au chaud vers 40 °C à 50 °C. Pour une plante donnée, on observe des modifications du point de compensation (IC) et du point de début de saturation (IS), mais sans modification du rendement Ф. Ceci montre que les réactions photochimiques sont peu ou pas sensibles à la température, au contraire des réactions biochimiques.
Plusieurs types d’expériences ont montré que la photosynthèse pouvait être découpée en deux groupes de réactions de significations différentes, mais couplées entre elles et nécessitant des intermédiaires.
7.1. Origine de l’oxygène (Ruben et Kamen, 1938)
L’équation bilan de la photosynthèse montre un dégagement de dioxygène.
La question se pose de savoir d’où provient ce dioxygène. On peut en effet émettre deux hypothèses : soit cet oxygène provient du CO2, soit il provient de l’eau H2O. Afin de trancher entre ces deux possibilités, Ruben et Kamen ont utilisé un isotope lourd de l’oxygène (18O) à la place de l’oxygène habituel (16O) et ils ont marqué ainsi diverses molécules (H2O, CO2). Lorsque de l’eau est marquée par le 18O (H218O), le dioxygène produit par la photosynthèse devient marqué ; ce n’est pas le cas lorsque le CO2 est marqué par le 18O. Ils en déduisent que c’est l’eau (H2O) qui est à l’origine du dioxygène produit. Pour former une molécule de dioxygène, il faut donc 2 molécules d’eau.
Ces résultats montrent que l’on peut décomposer la réaction photosynthétique en deux groupes de réactions :
Figure 23 – Équation bilan des deux groupes de réaction de la photosynthèse phase claire et phase sombreCes deux réactions (oxydation de l’eau et réduction du dioxyde de carbone) sont couplées dans un ensemble complexe de réactions d’oxydoréduction faisant intervenir des transporteurs de protons (H+) et d’électrons (e–).Auteur(s)/Autrice(s) : Roger Prat, François MoreauLicence : Pas de licence spécifique (droits par défaut)
7.2. Existence de deux types de réactions (Emerson et Arnold, 1932)
Diverses expériences d’incorporation de CO2 par des chlorelles (algues unicellulaires) ont permis de mettre en évidence que l’ensemble des réactions composant la photosynthèse peut être décomposé en deux groupes :
des réactions mettant directement en jeu la lumière – on parle de phase photochimique de la photosynthèse ;
des réactions plus lentes, sans utilisation directe de la lumière – on parle de phase biochimique de la photosynthèse.
7.3. La libération d’O2 nécessite un accepteur d’électron (Hill, 1937)
Hill utilise une suspension de chloroplastes isolés dans un tampon sans CO2. Il mesure les variations de dioxygène à l’aide d’une électrode à oxygène. Il ajoute à la préparation un accepteur artificiel d’électrons, le ferricyanure de potassium, Fe3+(CN–)6K3 (réactif de Hill) et travaille en lumière continue.
Figure 25 – Nécessité d’un absorbeur d’électron pour la photosynthèseEn absence de CO2, les chloroplastes sont capables de libérer du dioxygène, à condition qu’un accepteur d’électron (Fe3+) soit présent dans le milieu.Auteur(s)/Autrice(s) : Roger Prat, François MoreauLicence : Pas de licence spécifique (droits par défaut)
Le réactif de Hill se comporte comme un accepteur d’électron :
Fe3+ + e – → Fe2+
Dans les conditions naturelles de la photosynthèse, ce rôle d’accepteur d’électron est rempli par le couple NADP+/NADPH :
NADP+ + 2e – + 2H+ → NADPH + H+
Le couple NADP+/NADPH joue ainsi le rôle d’intermédiaire entre l’oxydation de l’eau et la réduction du CO2. De plus, l’ATP sert aussi d’intermédiaire énergétique : de l’ATP est formé en conséquence du fonctionnement de la chaîne photosynthétique, et est ensuite utilisé lors de la formation des composés carbonés.
7.4. Schéma bilan
Figure 26 – Représentation schématique statique de la séparation de la photosynthèse en deux groupes de réactionAuteur(s)/Autrice(s) : Roger Prat, François MoreauLicence : Pas de licence spécifique (droits par défaut)
La plupart d’entre nous sont familiarisée aux mots et aux concepts tels que changement climatique, emissions de dioxyde de carbone, pollution de l’air et pénurie d’eau, mais nous sommes très peu à avoir porté notre attention sur les sols. Pendant des millénaires, la vie sur terre a été soutenue par une mince couche de sols fertiles sur la croûte terrestre.
Mais… Que sont les sols et que font-il exactement ?














 décomposé par les bactéries et les champignons, l'évolution du dioxyde de carbone et les protozoaires et les nématodes consommant les bactéries et les champignons et excrétant de l'ammonium dans le sol pour la croissance des plantes. NO3- et NH4+ sont facilement convertis dans le sol. Graphique du Dr Rafiq Islam.")
 décomposé par les bactéries et les champignons, l'évolution du dioxyde de carbone et les protozoaires et les nématodes consommant les bactéries et les champignons et excrétant de l'ammonium dans le sol pour la croissance des plantes. NO3- et NH4+ sont facilement convertis dans le sol. Graphique du Dr Rafiq Islam.")